
A 2026 fMRI study of 79 women with interpersonal-violence-related PTSD found that the hippocampus does encode the meaning of autobiographical trauma narratives, t(140) = 4.05, p < 0.001, even though average hippocampal encoding did not rise for trauma vs. neutral memories.
Research Highlights
- Hippocampal semantic encoding was significant: Cisler et al. analyzed 3,928 narrative sentences in 79 women with PTSD and found that hippocampal activity tracked sentence meaning, t(140) = 4.05, p < 0.001.1
- Trauma and neutral memories did not differ by mean hippocampal encoding: the trauma-vs-neutral mean comparison was not significant, p = 0.27, which makes the result a pattern-level finding rather than a simple activation increase.1
- CA1 and dentate gyrus carried the trauma split: spatial semantic-encoding patterns distinguished trauma from neutral narratives in CA1, AUC = 0.69, p = 0.002, and dentate gyrus, AUC = 0.66, p = 0.01.1
- PTSD severity tracked hippocampal encoding: higher CAPS-5 symptom severity was associated with stronger hippocampal semantic encoding than lower PTSD severity, regardless of narrative type, t(140) = 3.56, p < 0.001.1
- The posterior cingulate moved in the opposite direction: semantic encoding was greater for neutral than trauma narratives in the PCC, t(147) = −2.95, p = 0.004, while left superior temporal gyrus encoding was significant overall, t(147) = 3.22, p = 0.002.1
The useful correction is narrow but important: earlier PTSD imaging could miss hippocampal involvement if it asked only whether a region became more active during trauma recall. Cisler et al. asked whether voxelwise activity patterns tracked the meaning of each sentence in a personal narrative, and that question produced a hippocampal signal that mean-activation approaches can average away.
Semantic encoding means predicting brain activity from the meaning of language. In this study, each sentence from a participant’s trauma or neutral autobiographical script was converted into a 4,096-feature sentence-transformer embedding, then used in ridge-regression models to predict fMRI activity across 63,789 gray-matter voxels.1
79 Women Heard Their Own Trauma and Neutral Narratives During fMRI
Cisler et al. enrolled 79 adult women aged 21–50 with current PTSD related to interpersonal violence. The sample came from 2 sites: University of Wisconsin, n = 41, and University of Texas at Austin, n = 38.
One additional participant was excluded for excessive head motion.1
Each participant worked with research staff to create 2 personal scripts: one about the index traumatic event and one about a minimally arousing neutral autobiographical event. Each script was constrained to 330–350 words, recorded as audio, and paired with the script text on screen during fMRI.
Participants heard the neutral script 4 times first, then the trauma script 4 times.
The fixed order is a real caveat. Because neutral always came before trauma, trauma-vs-neutral differences can be confounded by order, habituation, fatigue, or later-run arousal.
The study also lacked a negative but non-traumatic control memory, so it cannot prove that every contrast is trauma-specific rather than negative-valence-specific.
The technical pipeline was unusually fine-grained for PTSD trauma-recall imaging. Whisper software extracted sentence timing from the audio files.
A sentence transformer called NV-Embed-V2 generated one 4,096-dimensional semantic vector for each sentence. The final matrices contained 3,928 sentences by 63,789 voxelwise activity values and 3,928 sentences by 4,096 semantic features.1
The Hippocampus Encoded Sentence Meaning, more than Trauma Arousal
The main hippocampal result confirmed the paper’s hypothesis: mean hippocampal semantic encoding was significant across participants, t(140) = 4.05, p < 0.001. That means the model could predict held-out hippocampal activity from sentence meaning better than chance, using 10-fold cross-validation across participants.1
The key calibration is the p = 0.27 trauma-vs-neutral mean comparison. Trauma narratives did not produce higher average hippocampal semantic encoding than neutral narratives.
Instead, the hippocampus encoded meaning in both narrative types, while the spatial pattern of that encoding differed by narrative type.
This distinction explains why prior literature could look contradictory. Sartory et al. meta-analyzed symptom-provocation fMRI studies and helped establish the older problem: trauma recall did not yield a clean, reliable hippocampal activation signature across studies.2
Thome et al. later reviewed trauma-related autobiographical-memory imaging and reached a similarly cautious baseline: hippocampal findings during trauma recall were inconsistent rather than absent in any simple biological sense.3
Perl et al. sharpened the question in 2023 by comparing traumatic and sad autobiographical memories in PTSD, but Cisler et al. argue that word-level embeddings averaged into broad narrative summaries can still blur sentence-specific meaning.4 The 2026 study does not prove Perl et al. were globally wrong; it shows that a more granular sentence-level model detects hippocampal semantic encoding that broader methods can miss.
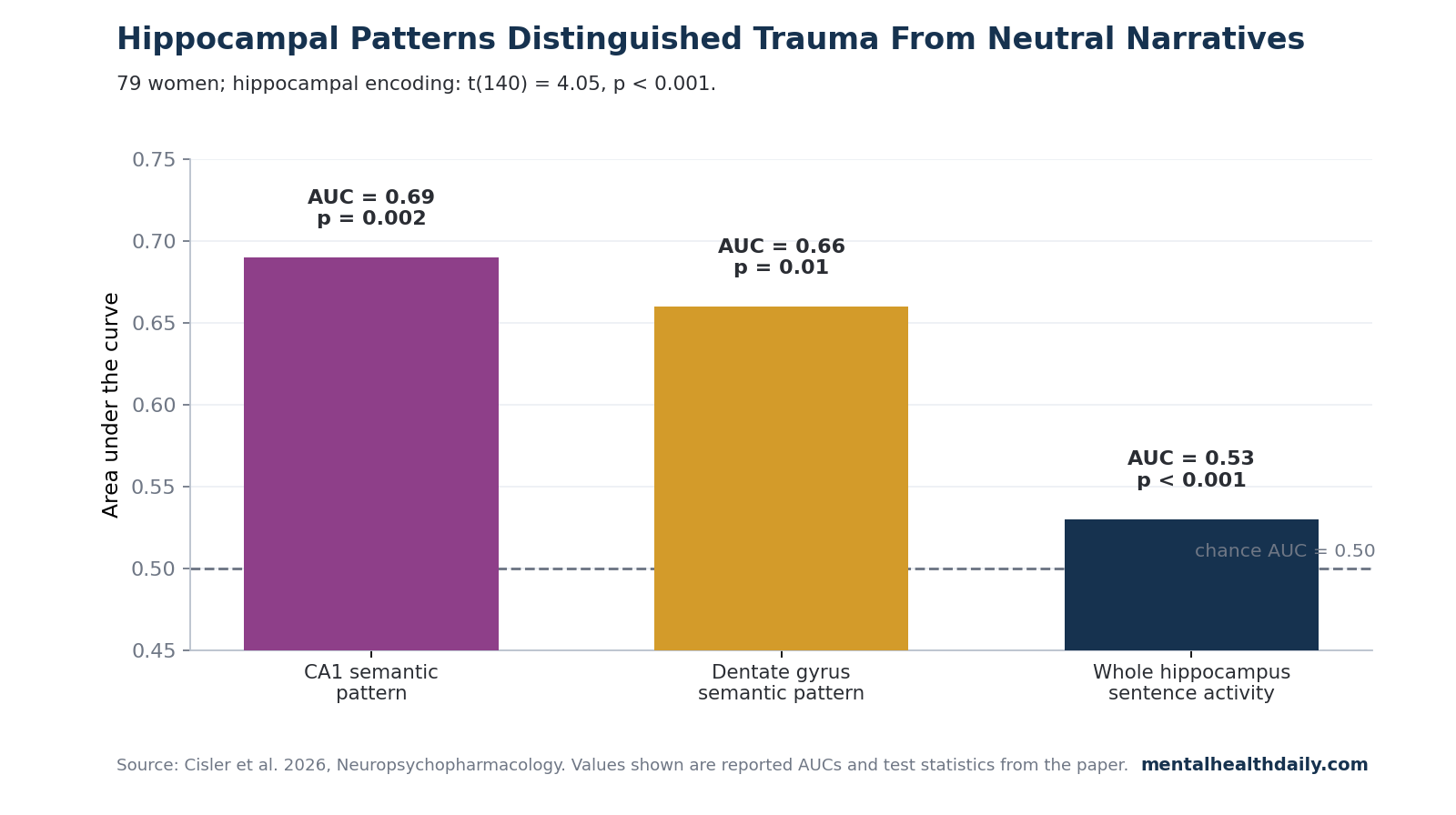
CA1 and Dentate Gyrus Separated Trauma From Neutral Narrative Patterns
CA1 is a hippocampal subfield involved in autobiographical memory and fear-extinction learning. Dentate gyrus is heavily tied to pattern separation, the process of keeping similar memories from collapsing into each other.
In PTSD, those functions matter because trauma cues can make past threat feel present, specific memories can generalize across superficially similar situations, and exposure therapy repeatedly asks patients to retrieve and re-contextualize a trauma narrative.
Cisler et al. found that spatial semantic-encoding patterns in CA1 differentiated trauma from neutral narratives with AUC = 0.69, p = 0.002. Dentate gyrus showed a similar but slightly smaller separation, AUC = 0.66, p = 0.01.
These are not large diagnostic-classifier effects; the paper describes the magnitudes as modest. They are still the most specific anatomical result in the study because they locate the trauma-vs-neutral pattern difference inside hippocampal subfields rather than in a generic “hippocampus involved” claim.1
Cross-narrative models pushed the same interpretation. Models trained on one narrative type did not generalize to the other narrative type in the hippocampus, with ps > 0.5.
If trauma and neutral memories used the same hippocampal semantic tuning, a model trained on neutral sentence meanings should have carried over more cleanly to trauma sentence meanings. It did not.
The result fits the clinical idea of trauma memories as complex narratives rather than single stored objects. A trauma memory contains “hot spots,” contextual details, sensory fragments, and lower-distress connective tissue.
Collapsing the full script into one average brain response is exactly the kind of analysis that can miss which parts of the memory are being encoded differently.
PTSD Severity Was Linked to Stronger Hippocampal Encoding
PTSD symptom severity was positively associated with hippocampal semantic encoding, t(140) = 3.56, p < 0.001, after adjustment for age, education, medication, scanner site, and framewise displacement. Subfield analyses found significant associations across hippocampal subfields, ts > 2.9 and ps < 0.009, except for the subiculum, p = 0.34.1
The interpretation is not that stronger hippocampal encoding causes PTSD. The study is cross-sectional, all participants already had PTSD, and symptom severity was measured above a diagnostic threshold.
A safer reading is that more severe PTSD was associated with stronger hippocampal tracking of autobiographical sentence meaning, across trauma and neutral scripts.
That association matters for treatment research because trauma-focused therapies such as prolonged exposure and cognitive processing therapy depend on repeated, structured contact with the trauma narrative. Cisler et al. do not show that therapy changes CA1 or dentate gyrus encoding.
They do make it harder to defend the older shortcut that the hippocampus is simply absent from trauma-memory recall.
The Posterior Cingulate Result Cuts Against a Simple Memory-System Story
The posterior cingulate cortex (PCC) is a major hub of the default mode network, a set of brain regions involved in self-directed thought, autobiographical memory, and internally oriented cognition. Cisler et al. found greater PCC semantic encoding for neutral than trauma narratives, t(147) = −2.95, p = 0.004.
Whole-brain results also pointed toward decreased trauma semantic encoding in medial prefrontal/default-mode regions.1
Left superior temporal gyrus (STG), a language-and-semantic-processing region, showed significant semantic encoding overall, t(147) = 3.22, p = 0.002. Cross-narrative STG models also generalized, t(147) = 2.49, p = 0.013.
Unlike the hippocampus, neither PCC nor STG encoding tracked PTSD symptom severity, with p = 0.55 for PCC and p = 0.46 for STG.
The positive pattern is specific: hippocampal patterns tracked sentence meaning and symptom severity; PCC/default-mode encoding was lower for trauma than neutral narratives; STG behaved more like a language-semantic region than a PTSD-severity marker.
Morris et al. reached a neighboring conclusion from a different decoding angle, showing that trauma recall in PTSD can be analyzed through threat-representation patterns rather than only broad region activation.5
What This Study Does and Does Not Show
Cisler et al. strengthen the hippocampal account of PTSD trauma recall, but the study should not be inflated into a brain-reading claim. The model predicted fMRI activity from known sentence meanings.
It did not reconstruct private memory content from brain activity, diagnose PTSD from a scan, or prove that CA1 and dentate gyrus store trauma memories as isolated modules.
- It supports hippocampal involvement: sentence meaning predicted hippocampal activity, t(140) = 4.05, p < 0.001.
- It supports pattern-level trauma specificity: CA1 and dentate gyrus semantic patterns distinguished trauma from neutral scripts, AUC = 0.69 and 0.66.
- It does not isolate trauma from negative emotion: the study did not include a negative non-trauma script.
- It does not generalize to all PTSD populations: the sample was limited to 79 women with interpersonal-violence-related PTSD.
The practical takeaway is methodological: PTSD trauma-memory imaging needs analyses that preserve sentence-level meaning, subfield-level patterning, and narrative heterogeneity. If a model averages the entire script into one response, it can erase the very memory structure it is trying to measure.
Questions About This PTSD Hippocampus Study
Did the study prove that trauma memories are stored in CA1 and dentate gyrus?
No. CA1 and dentate gyrus patterns distinguished trauma from neutral narratives, with AUC = 0.69 and 0.66, but that is pattern classification during recall, not proof of a storage location.
The better reading is that these subfields participate in how traumatic narrative meaning is represented during recall.
Why is p = 0.27 important if the hippocampal result was significant?
The p = 0.27 result means average hippocampal semantic encoding did not differ significantly between trauma and neutral narratives. The significant finding was that the hippocampus encoded meaning in both, and that the spatial pattern of encoding differed by narrative type.
That is a stronger and more precise claim than “trauma made the hippocampus more active.”
Does this overturn the 2023 traumatic-vs-sad memory paper?
It narrows the conclusion. Perl et al. found evidence that traumatic and sad autobiographical memories differ in neural representation, and Cisler et al. agree that trauma narratives have distinct patterns.
The disagreement is about the hippocampus: Cisler et al. show that sentence-level semantic modeling detects hippocampal encoding that broader word-averaged or script-averaged approaches can miss.
Can this fMRI method diagnose PTSD in one patient?
No. The models were cross-validated across participants to test group-level semantic encoding.
The paper does not report an individual diagnostic tool, and whole-brain symptom-severity models found no significant clusters after correction.
What would make the trauma-specific claim stronger?
A follow-up study would need randomized script order, a negative non-trauma autobiographical memory, trauma-exposed controls without PTSD, and a broader sample that includes men and other trauma types. Those design changes would separate trauma content from order, arousal, negative emotion, and interpersonal-violence-specific sampling.
References
- Semantic encoding of trauma memories in the hippocampus among individuals with PTSD. Cisler JM, Malloy LT, Jaeb M, Dunsmoor JE, Stowe ZN. Neuropsychopharmacology. 2026. doi:10.1038/s41386-026-02402-5
- In Search of the Trauma Memory: A Meta-Analysis of Functional Neuroimaging Studies of Symptom Provocation in Posttraumatic Stress Disorder (PTSD). Sartory G et al. PLOS ONE. 2013;8(3):e58150. doi:10.1371/journal.pone.0058150
- The neural correlates of trauma-related autobiographical memory in posttraumatic stress disorder: A meta-analysis. Thome J et al. Depression and Anxiety. 2020;37(4):321-345. doi:10.1002/da.22977
- Neural patterns differentiate traumatic from sad autobiographical memories in PTSD. Perl O et al. Nature Neuroscience. 2023;26:2226-2236. doi:10.1038/s41593-023-01483-5
- Decoding threat neurocircuitry representations during traumatic memory recall in PTSD. Morris KR et al. Neuropsychopharmacology. 2025;50:568-575. doi:10.1038/s41386-024-02028-5
- Prolonged exposure therapy for PTSD: emotional processing of traumatic experiences. Foa EB, Hembree EA, Rothbaum BO. Oxford University Press. 2007. doi:10.1093/med:psych/9780195308501.001.0001