
A 2025 Nature Communications study found that Alzheimer’s-like mice lost locus coeruleus noradrenergic axons in the olfactory bulb early: 14% loss by 2 months, 27% by 3 months, and 33% by 6 months, before broader forebrain axon degeneration appeared.1 Early smell loss may reflect a specific brainstem-to-olfactory circuit injury, while smell testing still needs biomarker and clinical context before it can inform Alzheimer’s risk.
Research Highlights
- Axon loss started early: AppNL-G-F mice showed 14% olfactory-bulb locus coeruleus axon loss at 2 months, rising to 27% at 3 months and 33% at 6 months.1
- Damage was region-specific first: Locus coeruleus axons in hippocampus, piriform cortex, and medial prefrontal cortex did not start degenerating until 6-12 months in the same model.1
- Smell loss had a circuit explanation: The study linked olfactory impairment to loss of noradrenergic input rather than only to late amyloid plaque burden.1
- Microglia were implicated: Olfactory-bulb microglia from 8 vs. 8 mice showed a phagocytosis-linked profile, and reducing phagocytosis preserved axons and olfaction.1
- Human data supported translation: Prodromal Alzheimer’s patients showed elevated olfactory-bulb TSPO-PET signal, and early Alzheimer’s post-mortem tissue showed reduced locus coeruleus axon density.1
Locus coeruleus means a small brainstem nucleus that supplies much of the brain’s noradrenaline, a neurotransmitter involved in arousal, attention, sleep-wake regulation, blood flow, memory, and sensory processing. In Alzheimer’s disease, the locus coeruleus is vulnerable early, but the clinical meaning of that early vulnerability has often been hard to pin down.
Olfactory dysfunction means impaired smell sensitivity or identification. It is common in Alzheimer’s disease and can precede clear cognitive decline, but smell loss also appears in Parkinson’s disease, viral illness, head injury, sinonasal disease, aging, smoking, and medication effects. That is why the circuit mechanism matters: it can make smell testing more biologically informed without pretending it is disease-specific.
Olfactory-Bulb Axon Loss Preceded Broader Forebrain Damage
Meyer et al. studied AppNL-G-F mice, an amyloid mouse model used to investigate Alzheimer’s-like pathology. At 1 month, olfactory-bulb locus coeruleus axon density was still unaltered compared with wild-type mice. By 2 months, axon density had fallen by 14%; by 3 months, by 27%; by 6 months, by 33%.1
Region specificity: the same early degeneration was not seen across all locus coeruleus projection fields. Hippocampus, piriform cortex, and medial prefrontal cortex showed later degeneration, beginning between 6 and 12 months.
Projection specificity: cholinergic and serotonergic projections in the olfactory bulb were also not reduced at 3 months, which made the signal more specific to the noradrenergic pathway.1
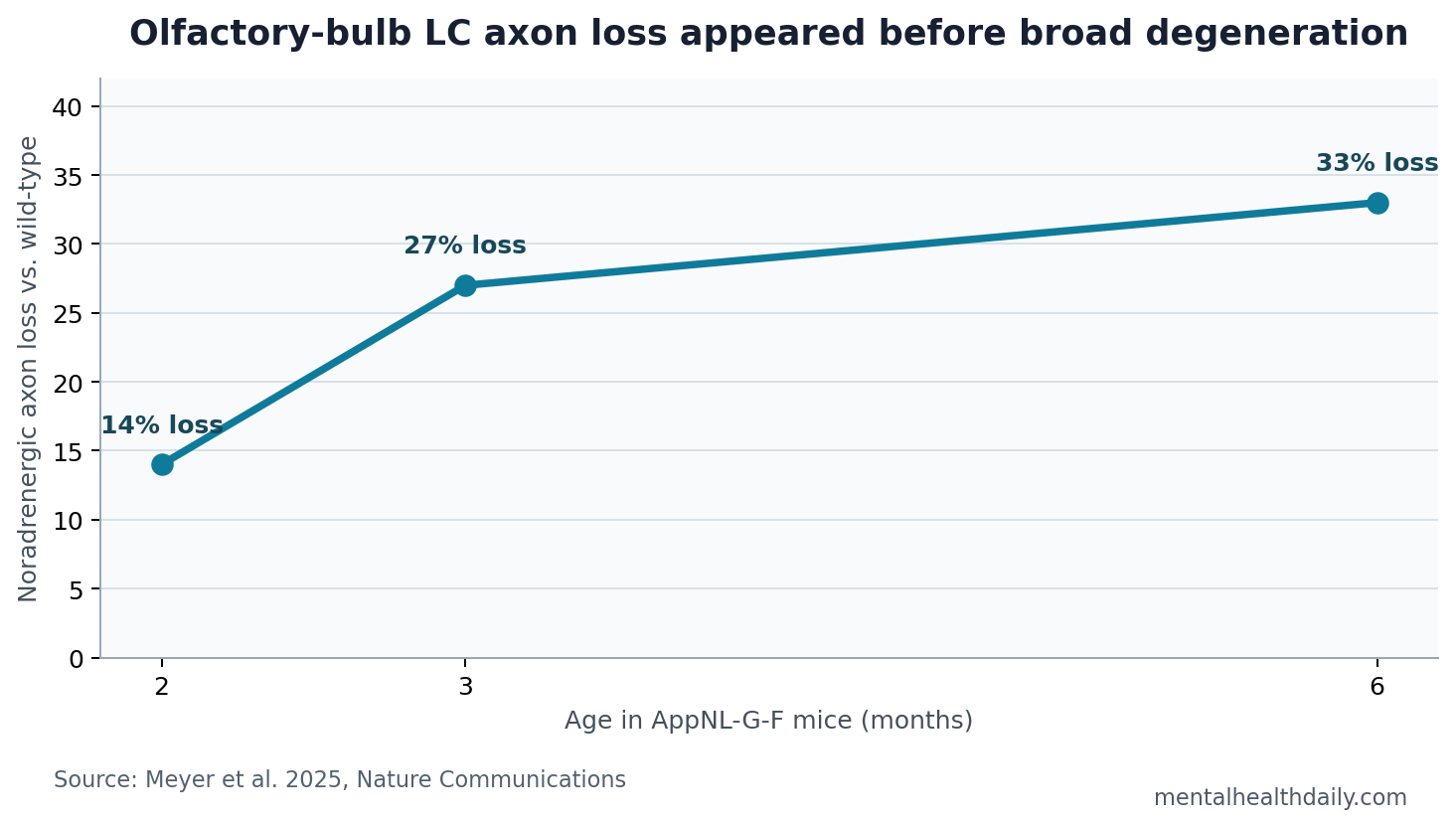
Plain-English interpretation: the smell pathway showed a targeted early projection injury rather than generalized late-stage brain degeneration. In this model, one projection from the locus coeruleus to the olfactory bulb looked unusually vulnerable.
The behavioral data were important because the axon finding could otherwise be dismissed as a microscope-only result. Meyer et al. used a buried-food task and an odor-discrimination task to show that AppNL-G-F mice had impaired olfactory performance when locus coeruleus input to the olfactory bulb was already reduced.1
Noradrenaline release also fell: with 2-photon imaging over the olfactory bulb, the researchers measured odor-stimulated noradrenaline dynamics during vanilla-puff exposure. AppNL-G-F mice showed reduced stimulus-linked noradrenaline release, which tied the structural loss of locus coeruleus fibers to a functional change in the smell circuit rather than only to reduced staining density.1
Microglia May Prune Noradrenergic Fibers Before Plaques Dominate
Microglia are resident immune cells in the brain. They clear debris, sculpt synapses, respond to injury, and can remove neural structures when those structures display “eat me” signals. In the Meyer et al. study, olfactory-bulb microglia appeared to recognize and phagocytose locus coeruleus axons.1
Phagocytosis means cellular engulfment. The paper’s mechanism centered on microglial recognition of externalized phosphatidylserine on locus coeruleus axons. When researchers reduced phagocytosis genetically, locus coeruleus axons and olfaction were preserved in the mouse model.1
Several details made that pruning interpretation more concrete. Olfactory-bulb microglia from 2-month-old AppNL-G-F mice showed a phagocytosis-linked transcript profile in bulk sequencing, and primary olfactory-bulb microglia from the model showed 33% higher phagocytic activity in vitro. Tissue imaging then placed locus coeruleus axon material inside microglia, connecting the gene-expression signal, cell behavior, and local axon loss in the same anatomical system.1
That mechanism is narrower and more useful than saying “inflammation causes smell loss.” The proposed chain is: amyloid-model biology changes olfactory-bulb microglia, vulnerable locus coeruleus axons display removal signals, microglia remove those axons, noradrenergic input drops, and olfactory behavior suffers.
TSPO-PET and Post-Mortem Tissue Put the Mouse Finding Near Human Disease
TSPO-PET is positron-emission tomography using a tracer for translocator protein, often interpreted as a marker related to glial activation. It is not a pure microglia meter, but it can show where inflammatory or glial signals are elevated in living people.
Meyer et al. reported elevated TSPO-PET signal in the olfactory bulb of prodromal Alzheimer’s patients, similar to the mouse-model olfactory-bulb gliosis signal. They also found decreased locus coeruleus axon density in post-mortem olfactory bulbs from patients with early Alzheimer’s disease.1
The human data do not prove that the mouse sequence is identical in every patient. They do make the mouse mechanism harder to dismiss as a local model artifact. The same anatomical zone showed a compatible glial signal in prodromal disease and compatible axon loss in early disease tissue.
Human limitation: the TSPO-PET cohort was small, with 16 patients in the prodromal Alzheimer’s group, and TSPO signal is not specific to one microglial state. The study therefore supports a translational bridge, not a clinical rule that olfactory-bulb TSPO-PET should be ordered for routine Alzheimer’s screening.
Smell Loss Is a Triage Signal, Not an Alzheimer’s Diagnosis
Olfactory impairment has long been discussed as an early Alzheimer’s marker. Attems et al. reviewed evidence that olfactory dysfunction and olfactory-system pathology appear in Alzheimer’s disease, but they also emphasized that the olfactory system is vulnerable across multiple neurodegenerative and non-neurodegenerative conditions.2
Braak and Del Tredici placed early Alzheimer’s-related pathology near brainstem systems including the locus coeruleus, which gave the field a staging reason to care about this nucleus before dementia is clinically obvious.3 Meyer et al. add a sensory-circuit reason: early locus coeruleus injury may be read out through smell before broader cognitive networks collapse.
Clinical implication: smell testing is most useful as part of a risk-stratification bundle. A low smell-identification score is more informative when paired with age, cognitive screening, family history, plasma or cerebrospinal-fluid biomarkers, sleep and Parkinsonian symptoms, sinonasal history, and medication review.
Noradrenaline Links Sensory Processing, Attention, and Alzheimer’s Vulnerability
Noradrenaline is the main neurotransmitter released by locus coeruleus neurons. It helps tune signal-to-noise in sensory systems, regulates arousal, and modulates immune and vascular responses. Losing noradrenergic tone can therefore affect more than one symptom domain.
Heneka et al. and related model work have argued that locus coeruleus noradrenaline can restrain inflammatory and Alzheimer’s-like pathology.4 Meyer et al. fit that larger literature but make it more anatomically specific: the olfactory bulb may be an early site where noradrenergic axons are lost and microglia become functionally important.
That specificity is useful for early-detection research. Instead of treating smell loss as a black-box symptom, future studies can ask whether olfactory-bulb TSPO-PET, locus coeruleus imaging, plasma p-tau217, amyloid status, and smell identification jointly predict cognitive decline better than any one marker alone.
What This Study Can and Cannot Support
Supported: in an Alzheimer’s amyloid mouse model, locus coeruleus axons into the olfactory bulb degenerated early and specifically; olfactory behavior declined alongside that loss; microglial phagocytosis looked mechanistically important; human prodromal and post-mortem data supported olfactory-bulb involvement.
Not supported: smell loss alone diagnosing Alzheimer’s disease, microglia being globally harmful, or a claim that preserving locus coeruleus axons will prevent human dementia. The causal rescue experiment was preclinical, and the human data were supportive rather than interventional.
Best next test: longitudinal human studies should combine smell testing with Alzheimer’s blood biomarkers, locus coeruleus-sensitive MRI, olfactory-bulb PET or MRI measures, and cognitive follow-up. The central question is whether this circuit signal improves prediction beyond cheaper and broader risk markers.
Questions About Smell Loss and Alzheimer’s Risk
Does poor smell mean someone has Alzheimer’s disease?
No. Smell loss has many causes. It becomes more concerning when it appears with cognitive change, biomarker evidence, rapid decline, REM sleep behavior disorder, Parkinsonian symptoms, or other neurodegenerative signs.
Why would the locus coeruleus affect smell?
The locus coeruleus sends noradrenergic fibers to the olfactory bulb. Those fibers help modulate sensory processing, so losing them can impair odor handling before a person has obvious memory symptoms.
What makes the Meyer et al. result stronger than a simple association?
The study combined time-course mouse data, microglial mechanism experiments, olfactory behavior, prodromal human PET, and post-mortem human tissue. That multi-layer design is stronger than a single smell-test correlation.
What should readers not take from this?
Do not treat smell testing as a standalone Alzheimer’s screen. It is a triage clue that needs biological context.
References
- Meyer C, Niedermeier T, Feyen PLC, et al. Early locus coeruleus noradrenergic axon loss drives olfactory dysfunction in Alzheimer’s disease. Nature Communications. 2025. doi:10.1038/s41467-025-62500-8
- Attems J, Walker L, Jellinger KA. Olfaction and aging: a mini-review. Gerontology. 2015. doi:10.1159/000381619
- Braak H, Del Tredici K. The pathological process underlying Alzheimer’s disease in individuals under 30. Acta Neuropathologica. 2011. doi:10.1007/s00401-010-0789-4
- Heneka MT, Nadrigny F, Regen T, et al. Locus ceruleus controls Alzheimer’s disease pathology by modulating microglial functions through norepinephrine. Proceedings of the National Academy of Sciences. 2010. doi:10.1073/pnas.0909586107