
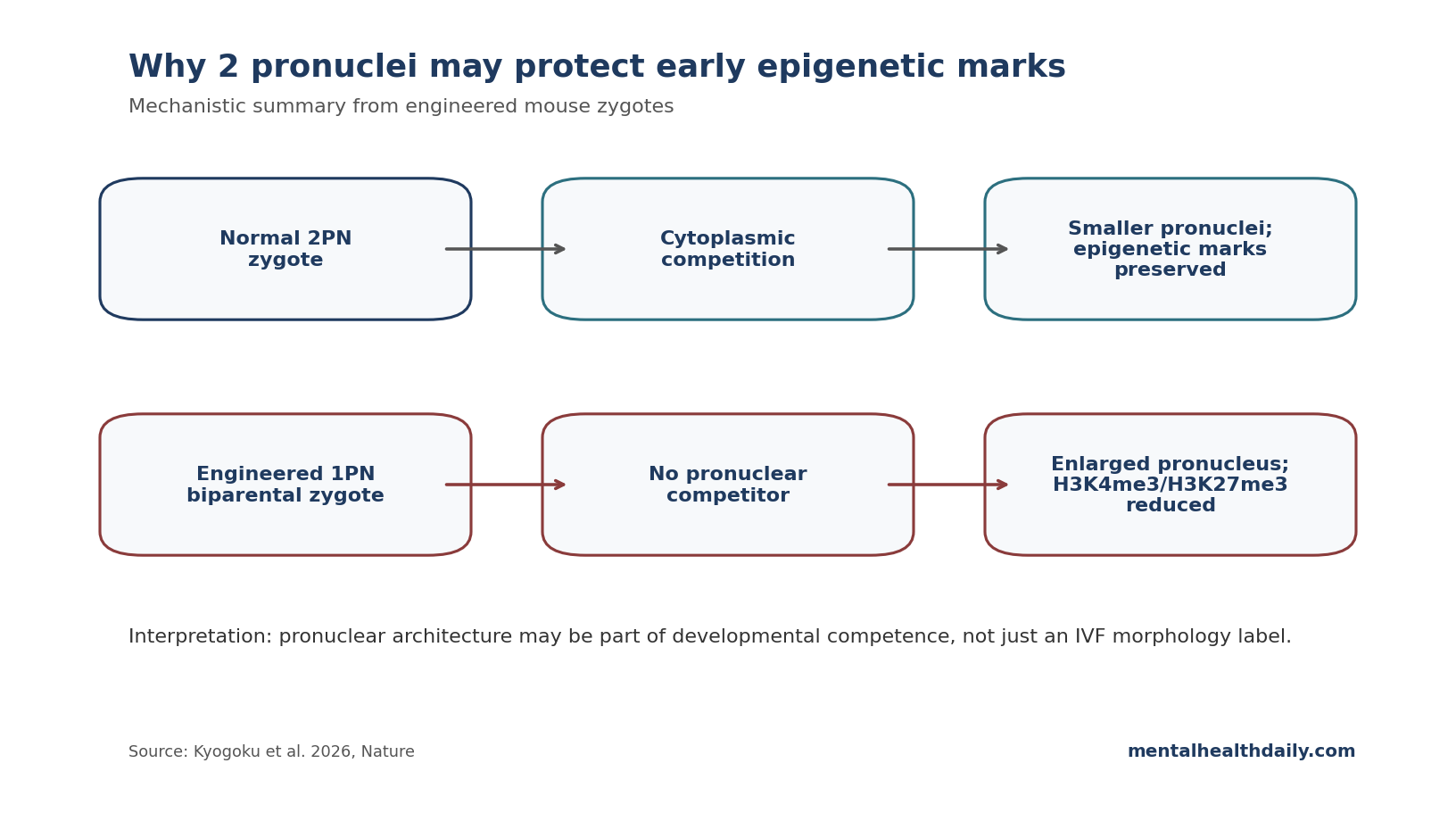
One-pronuclear biparental zygotes may be less biologically normal than they look. In a 2026 Nature mouse study, forcing maternal and paternal genomes into 1 enlarged pronucleus disrupted early embryonic epigenetic marks, while the usual 2-pronuclear state created a cytoplasmic competition that helped preserve developmental potential.1
Research Highlights
- The 2026 zygote study asks why mammalian zygotes start with 2 pronuclei. Maternal and paternal chromosomes normally sit in separate pronuclei until the first mitosis, even though most later cells use 1 nucleus.1
- One-pronuclear biparental zygotes formed an enlarged pronucleus. The enlarged structure contained spatially separated parental chromosome regions, but key histone marks were reduced.1
- H3K4me3 and H3K27me3 were the main warning signals. These trimethylated histone marks help organize early developmental chromatin; they were lower in 1PN biparental zygotes.1
- The proposed mechanism is cytoplasmic competition. Separate pronuclei compete for cytoplasmic material, limiting pronuclear volume; without that competition, pronuclear enlargement appears to dilute or destabilize epigenetic regulation.1
- The clinical caveat is calibrated embryo selection. Some human 1PN biparental zygotes can develop, but the mouse data argue that “biparental diploid” still leaves epigenetic uncertainty compared with normal 2PN development.1
The compact version: 2 pronuclei may help set the physical conditions required for early epigenetic programming, especially by preventing 1 giant pronucleus from forming before the embryo has stabilized its chromatin state.
1PN Biparental Zygotes Are Clinically Tempting but Biologically Ambiguous
In mammalian fertilization, the maternal and paternal genomes initially sit in separate pronuclei. This 2-pronuclear, or 2PN, stage is visually familiar in IVF and ICSI workflows. One-pronuclear, or 1PN, zygotes are also seen in assisted reproduction. Some are abnormal, but some are biparental diploids, meaning they contain both maternal and paternal chromosome sets and can sometimes develop.1
That creates a practical question. If a 1PN zygote is biparental and diploid, is it merely a different-looking normal embryo, or does the single-pronucleus state itself carry biological cost? Kyogoku et al. built a mouse system to test that question directly.1
A compact study snapshot:
- Model: mouse zygotes generated with ICSI-based manipulation.
- Main comparison: normal 2PN zygotes versus engineered 1PN biparental zygotes.
- Key measurements: pronuclear volume, parental chromosome separation, histone marks, genome-wide H3K4me3/H3K27me3 profiles, live imaging, and development to term.
- Core mechanism tested: whether separate pronuclei compete for shared cytoplasmic resources and thereby limit pronuclear growth.
The paper leaves room for clinically observed 1PN human embryos that develop, while arguing that pronuclear number may carry mechanistic information beyond descriptive morphology.
1PN Zygotes Kept Parental Regions Apart but Lost Histone Marks
The engineered 1PN biparental zygotes formed a large pronucleus whose volume was close to the combined volume of the maternal and paternal pronuclei in 2PN controls. Inside that enlarged pronucleus, maternal and paternal chromosome regions remained partly distinct, marked by 5-methylcytosine and 5-hydroxymethylcytosine patterns.1
That is the subtle point. Maternal and paternal chromosomal territories were still spatially organized. The defect appeared in epigenetic marks, especially H3K4me3 and H3K27me3.
H3K4me3 and H3K27me3 are histone modifications, chemical tags on chromatin proteins that help regulate gene activity and developmental timing. In early embryos, these marks are not decorative. They are part of the genome's transition from gamete state to embryonic state.2,4
In 1PN biparental zygotes, total H3K4me3 and H3K27me3 signals were reduced. More focused maternal-region measurements showed significantly lower H3K4me3 and H3K27me3 in both the nuclear interior and peri-nucleolus precursor-body regions. H3K27ac was also reduced, suggesting a broader disturbance in chromatin regulation.1
Cytoplasmic Competition Limited Pronuclear Volume
Volume control: the mechanism is surprisingly physical. Separate pronuclei appear to compete for shared cytoplasmic material. When there are 2 pronuclei, neither grows unchecked. When the embryo contains only 1 enlarged biparental pronucleus, that competitive volume control is lost.1
Model test: the researchers tested this through halved and doubled zygote experiments, live pronuclear-growth imaging, and a theoretical model. Pronuclear volume scaled with cytoplasmic volume, and the growth kinetics matched the model's prediction that pronuclei compete through the cytoplasm.1
Plain English: early embryos may need 2 smaller nuclear compartments because the cytoplasm can only supply so much nuclear-building and chromatin-maintenance material. If all the parental DNA sits in 1 large compartment, the local nuclear environment changes enough to disturb epigenetic marks.
That is a different way to think about early development. The usual mental model is gene-centered: does the embryo have the right chromosomes and the right parental origin? This paper adds a geometry-centered layer. The same genetic material can sit inside a nuclear compartment of the wrong size, and that physical arrangement can change the chemical landscape around chromatin.
The finding also helps explain why the 2PN state might persist despite carrying a cost. Separate pronuclei can create chromosome-segregation risks at the first mitosis because the maternal and paternal chromosome sets start apart. If evolution kept this arrangement anyway, it likely supplies a compensating benefit. The proposed benefit here is early nuclear-volume control and preservation of chromatin marks before the first embryonic divisions.
Rescue Experiments Strengthened the Mechanism
The strongest part of the paper is that the researchers did not stop at correlation. If pronuclear enlargement causes epigenetic loss, then suppressing enlargement should rescue at least part of the defect. They tested this by transiently introducing an extra pronucleus as a cytoplasmic competitor, then removing it after pronuclear growth.1
That competition-based rescue reduced the volume of the biparental pronucleus and strongly rescued maternal H3K4me3 and H3K27me3 levels. Genome-wide profiling also showed recovery of paternal promoter H3K4me3 enrichment, although paternal H3K27me3 remained less rescued, suggesting more than 1 mechanism may be involved.1
The researchers also tested drug-based restoration of epigenetic marks as a mechanistic proof. If epigenetic-mark loss is a key mediator of the 1PN problem, then restoring those marks should improve developmental potential.
Developmental Potential Was Reduced, Not Absent
Developmental signal: the clinical nuance is developmental potential, not deterministic failure. The paper does not claim that 1PN biparental zygotes can never develop. In human assisted reproduction, some 1PN biparental zygotes have produced apparently healthy births, which is why they have been considered as possible embryos for use when options are limited.1
The mouse data sharpen the concern. A 1PN biparental state can be chromosomally plausible while still being epigenetically compromised. “Both parental genomes are present” is not the same as “the early nuclear architecture did the same developmental work.”
This fits broader preimplantation epigenetics. Human and mouse embryos undergo extensive DNA methylation and histone-mark reprogramming after fertilization.2,3 Small changes in timing, compartment size, or chromatin-state establishment can matter disproportionately because the embryo is setting the initial regulatory conditions for every later lineage.
Clinical translation: the paper's most practical contribution is a warning against reducing embryo competence to a few visible or chromosomal features. A 1PN embryo might pass some genetic checks and still have unresolved epigenetic uncertainty. Conversely, the existence of successful 1PN-derived births means the finding supports calibrated risk assessment instead of a universal ban or deterministic risk claim.
The honest clinical posture is calibrated: normal 2PN development remains the cleaner biological default; 1PN biparental embryos may sometimes be usable; and this mouse work argues that 1PN use should be studied with epigenetic, developmental, and long-term follow-up endpoints rather than morphology alone.
How to Interpret 1PN Zygotes in IVF Context
For embryo biology: pronuclear number may reflect whether the embryo has the correct physical setup for epigenetic regulation.
For IVF interpretation: a biparental diploid 1PN zygote may be better than a clearly abnormal embryo, but it should not automatically be considered equivalent to a normal 2PN zygote.
For translation: this is a mouse study with engineered 1PN zygotes. Human clinical use requires human embryo data, careful follow-up, and context-specific risk-benefit thinking.
For the broader point: developmental competence includes chromosome count, nuclear architecture, cytoplasmic resource allocation, and epigenetic marks.
Human translation should therefore focus on prospective evidence rather than isolated success stories. A live birth after 1PN transfer proves possibility, but developmental policy needs denominator data, embryo-genetics context, pregnancy outcomes, and child follow-up when ethically available.
Embryology labs also need clear language for uncertainty. Calling a 1PN embryo normal because chromosomes look biparental can understate nuclear-architecture and epigenetic questions that this mouse work raises.
That language matters most when patients are weighing limited embryo options.
Clear uncertainty is better than false equivalence.
Questions About One-Pronuclear Zygotes
Why does a 1PN zygote raise an embryo-selection question?
A 1PN zygote has 1 visible pronucleus instead of the usual separate maternal and paternal pronuclei seen after fertilization.
Did the mouse 1PN zygotes contain both parental genomes?
Yes. The study engineered biparental 1PN zygotes and showed maternal and paternal chromosome regions remained partly separated inside the enlarged pronucleus.1
What went wrong?
The enlarged single pronucleus showed reduced H3K4me3, H3K27me3, and H3K27ac signals, pointing to disrupted early epigenetic regulation.1
Does this settle human IVF practice?
No. It provides a mechanistic mouse warning. Clinical embryo-use decisions require human evidence, embryo-specific context, and specialist reproductive-medicine oversight.
References
- Cytoplasmic Competition Between Separate Parental Pronuclei in Zygotes. Kyogoku H, Tarama M, Matsuwaka M, et al. Nature. 2026. doi:10.1038/s41586-026-10417-7
- DNA Methylation Dynamics of the Human Preimplantation Embryo. Smith ZD, Chan MM, Humm KC, et al. Nature. 2014;511:611–615. doi:10.1038/nature13581
- Chromatin Dynamics in the Regulation of Cell Fate Allocation During Early Embryogenesis. Burton A, Torres-Padilla ME. Nature Reviews Molecular Cell Biology. 2014;15:723–734. doi:10.1038/nrm3885
- Allelic Reprogramming of the Histone Modification H3K4me3 in Early Mammalian Development. Zhang B, et al. Nature. 2016;537:553–557. doi:10.1038/nature19361