
A 2026 Nature Communications mouse study tested 7,736 possible connections in the hippocampal CA3 memory circuit and found a developmental shift from dense early wiring toward sparse structured adult wiring: corrected CA3-to-CA3 connection probability fell from 6.37% at P7-8 to 2.17% at P45-50.1 The finding supports a circuit-level version of memory maturation, not a claim about boosting human memory with a simple intervention.
Research Highlights
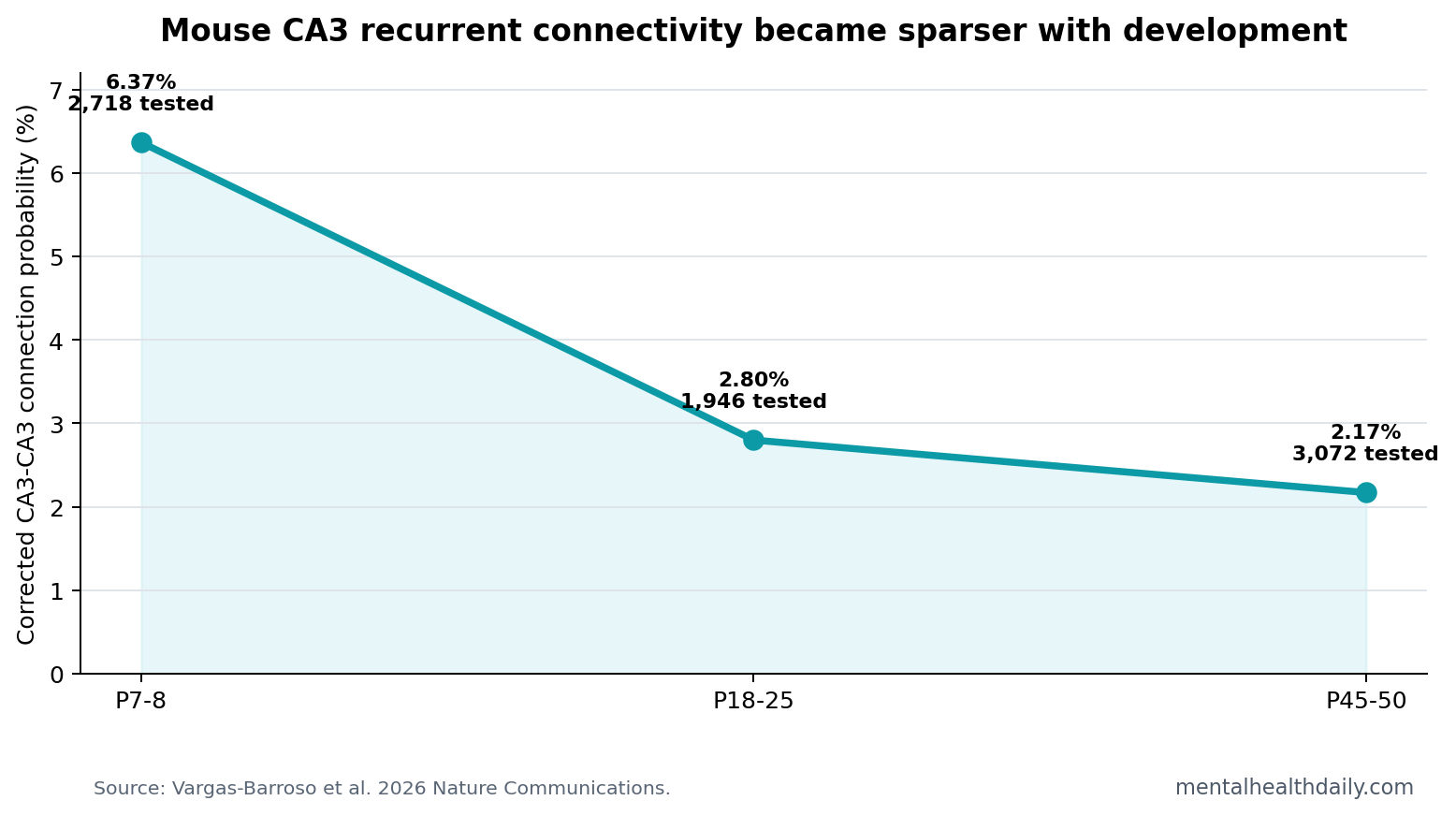
- 7,736 connections were tested: recordings covered 2,718 possible connections at P7-8, 1,946 at P18-25, and 3,072 at P45-50.1
- Connectivity fell with age: corrected CA3-CA3 connection probability declined from 6.37% to 2.8% to 2.17% across the 3 developmental windows.1
- The adult circuit was more structured: the network moved from local, dense, and random connectivity toward distributed, sparse, and structured organization.1
- Single synapses lost trigger power: early single synaptic events could drive postsynaptic spiking, while later activity required spatial summation of several inputs.1
- Translation is basic neuroscience: the data explain mouse CA3 development, not human memory testing or dementia treatment.
CA3 is a hippocampal subregion whose pyramidal neurons connect back onto one another. That recurrent architecture makes CA3 a candidate autoassociative network: a circuit that can store partial patterns and later complete them during memory retrieval.
Pattern completion means rebuilding a full memory representation from partial input. In theory, CA3 recurrent wiring is useful because one part of a stored representation can reactivate the rest. The unresolved question was whether the adult sparse structure starts as a prewired pattern or emerges during development.
CA3 Connectivity Dropped Across 3 Developmental Windows
Vargas-Barroso et al. used simultaneous multicellular patch-clamp recordings from mouse CA3 pyramidal neurons in acute hippocampal slices. The researchers stimulated putative presynaptic neurons with 5 action potentials at 20 Hz and recorded excitatory postsynaptic potentials and currents in possible postsynaptic partners.1
Patch clamp is an electrophysiology technique that records electrical activity from individual cells. Here, it let the researchers ask whether one CA3 pyramidal neuron actually drove another, with cell proximity alone kept out of the connectivity definition.
Measured connection probability fell from 3.38% at P7-8 to 1.64% at P18-25 and 0.91% at P45-50. After correcting for preserved axons, the same trajectory remained: 6.37%, 2.8%, and 2.17%, respectively. The early-to-juvenile comparison was significant at p = 0.002, and the early-to-adult comparison was p < 0.0001.1
Sparser Connectivity Became More Structured
The network became more distributed and structured as connection probability fell. Early CA3 wiring was more local and dense. Later wiring had broader axonal organization and more structured connectivity, consistent with an adult circuit designed for selective ensemble activation rather than indiscriminate excitation.
Functional implication: a dense immature network may be easier to ignite, but a sparse structured network can separate stored patterns more efficiently. That is the computational link to memory: useful retrieval needs both completion and discrimination.
Single Synapses Became Less Dominating
The paper also reported that synaptic transmission changed with development. Early in development, single synaptic events could be strong enough to trigger postsynaptic spiking. Later, individual synapses were weaker, and several inputs had to sum spatially before the postsynaptic cell crossed threshold.1
- Early circuit: dense local wiring plus strong synapses favors broad activation.
- Adult circuit: sparse structured wiring plus weaker individual synapses favors selective ensemble recruitment.
- Memory relevance: selective recruitment is closer to what pattern-completion models need for stable retrieval without runaway activation.
Animal-Slice Limits Are Central
Evidence-strength note: this was mouse ex vivo circuit physiology. It supports a developmental mechanism for CA3 wiring, not a human clinical conclusion. Acute slices remove intact behavioral state, neuromodulatory context, sleep-wake cycling, and the full hippocampal-entorhinal system.
The study is still valuable because it measures the circuit variable directly. Human imaging can infer hippocampal involvement in memory, and animal behavior can test memory performance, but multicellular recordings expose the wiring rule itself.
Why CA3 Needs Both Sparsity and Structure
A memory circuit that connects everything to everything would be powerful but unstable. Too much recurrent excitation makes stored patterns interfere with one another. Too little recurrent excitation makes pattern completion weak because partial cues cannot reactivate the rest of the ensemble.
The Vargas-Barroso result lands between those extremes. Connection probability fell across development, but the network gained structure. That combination is exactly what an autoassociative memory system needs: enough recurrent wiring to retrieve patterns, with enough selectivity to keep one memory from bleeding into another.
Early developmental logic: dense local wiring may help immature circuits establish activity and coordinate growth. Strong single synapses can recruit postsynaptic cells quickly, which may support early network formation before memory load becomes complex.
Adult computational logic: sparse structured wiring can support more specific ensemble activation. A partial cue can recruit the right pattern if recurrent inputs converge on a trained ensemble, while unrelated cells remain below threshold.
Developmental Timing Fits the Memory-Load Question
The 3 sampled windows matter. P7-8 mice are early postnatal animals. P18-25 mice are juvenile. P45-50 mice are closer to mature young adulthood. Across that trajectory, the animal’s sensory experience, exploratory behavior, and hippocampal memory demands change sharply.
The study does not prove that experience alone caused the wiring shift. Genetic programs, activity-dependent plasticity, cell maturation, axon growth, and pruning can all contribute. The important point is the direction: the mature CA3 circuit looked less like a dense prewired mesh and more like a sparse structured network.
Experience-dependent possibility: if repeated activity strengthens some recurrent routes and weakens others, network motifs could gradually reflect stored information. That would make adult CA3 structure partly a history of prior circuit use.
Preconfigured possibility: if developmental programs set the wiring trajectory in advance, sparse structure could emerge even without specific memory storage. The present data show the trajectory but do not fully separate those mechanisms.
Human Memory Claims Need a Translation Step
The article’s clinical relevance is conceptual. Human hippocampal disorders such as Alzheimer’s disease, temporal-lobe epilepsy, traumatic brain injury, and developmental neuropsychiatric conditions can involve memory networks. A mouse CA3 slice study helps explain what a mature memory circuit might require, but it does not diagnose those conditions or specify treatment.
For human research, the useful bridge would be cross-scale evidence: cellular wiring principles in animals, hippocampal subfield imaging in humans, memory-task behavior, and disease-specific pathology. Without that bridge, “memory circuit” language can sound more clinical than the evidence allows.
The Result Calibrates “Plasticity” Language
Plasticity boundary: developmental change does not automatically prove experience wrote the circuit. The CA3 data are more precise.
Connectivity probability, axonal organization, synaptic strength, and model behavior changed together across postnatal time. That is a real developmental transformation, with possible contributions from activity-dependent change, programmed maturation, pruning, and structural growth.
Useful interpretation: the mature memory circuit appears tuned and selective. Fewer connections can coexist with better information routing if the remaining connections are patterned. That is why the paper’s central value is the combined shift toward sparse structured connectivity and weaker single-synapse dominance.
This also helps explain why dense early circuitry should not be romanticized as “more connected” in a good sense. Developing brains need broad coordination, but mature memory systems need selectivity. More wiring is not automatically better wiring.
The same caution applies to weak synapses. A weaker single connection can look like loss if it is read in isolation. In an ensemble circuit, weaker individual inputs may be exactly what allows several converging cues to decide whether a cell joins the active memory pattern. The adult CA3 network appears to trade single-synapse dominance for multi-input integration.
That tradeoff gives the finding its cognitive relevance. Memory retrieval depends on partial cues recruiting the right ensemble while keeping competing ensembles quiet. Sparse structured connectivity is one plausible cellular route to that balance.
It also keeps the result grounded. The paper did not measure memory performance in the same animals, and it did not manipulate experience to see whether the same CA3 wiring pattern changed.
Its contribution is narrower and still valuable: it shows that the recurrent CA3 substrate available for memory computation changes markedly between early postnatal, juvenile, and mature windows. That gives later behavioral and disease studies a concrete circuit feature to test instead of a vague claim about hippocampal plasticity.
Questions About CA3 Memory Circuit Development
Does this mean childhood experience wires memory circuits?
Partly, but the study does not isolate experience from genetically guided development. It shows that CA3 recurrent connectivity becomes sparser and more structured across postnatal development.
Is lower connection probability bad?
No. In this circuit, lower probability with more structure may be adaptive because memory retrieval needs selective reactivation, not maximal excitation.
Does this apply to Alzheimer’s disease?
Only indirectly. CA3 is relevant to memory, but this study did not test dementia, aging, amyloid, tau, or human patients.
Why focus on CA3 instead of the whole hippocampus?
CA3 has recurrent pyramidal-neuron connections that make it a key candidate circuit for pattern completion. The study focused there because changing recurrent wiring is directly relevant to how partial cues may reactivate stored memory ensembles.
References
- Vargas-Barroso V, Watson JF, Navas-Olive A, Schlogl A, Jonas P. Developmental emergence of sparse and structured synaptic connectivity in the hippocampal CA3 memory circuit. Nature Communications. 2026. doi:10.1038/s41467-026-71914-x
- Marr D. Simple memory: a theory for archicortex. PubMed
- Rolls ET. Pattern separation, completion, and categorisation in the hippocampus and neocortex. PubMed
- Guzman SJ, et al. Synaptic mechanisms of pattern completion in the hippocampal CA3 network. PubMed