
A 2026 PNAS study found that TMEM175 behaves as a pH-sensitive lysosomal channel relevant to Parkinson’s risk biology.1 Acidifying the luminal side from pH 7.4 to 4.7 activated proton conductance, shifted reversal potential, and tied H57 to H+ and K+ flux.
Research Highlights
- Acidification activated TMEM175: lowering luminal-side pH from 7.4 to 4.7 increased inward and outward currents in TMEM175-expressing cells (n = 26), but not control cells (n = 11).
- The proton signal was transient: Erev shifted positive near +20 mV before returning toward 0 mV, supporting proton-gradient collapse rather than a stable pure H+ channel readout.
- Buffering changed the peak: 50 mM citrate produced a higher Erev peak than 5 mM acetate (+42.9 ± 6.9 mV, n = 27 vs. +25.6 ± 5.4 mV, n = 18).
- H57 was mechanistically important: the H57Y mutant reduced H+ and K+ conductance and altered H+/K+ selectivity in electrophysiological analyses.
- Clinical claims remain indirect: TMEM175 is linked to Parkinson’s risk biology, but this experiment did not test patients, symptoms, or disease-modifying treatment.
TMEM175 is a lysosomal cation channel. Lysosomes are acidic recycling organelles that help break down damaged proteins, worn-out cellular parts, and other cargo. In Parkinson’s disease, lysosomal failure matters because protein clearance and α-synuclein handling sit close to the disease mechanism.
Reversal potential, or Erev, is the voltage where net ion current through a channel reverses direction. For a proton-conducting channel, Erev should move with the proton gradient. Schulze et al. used that logic to ask whether TMEM175 current under acidic conditions is really H+ flow, K+ flow, or a mixed current distorted by rapid proton-gradient collapse.
pH 7.4 to 4.7 Shifted TMEM175 Toward Proton Conductance
TMEM175 had long been described as a lysosomal K+ channel, but prior acidic-pH experiments made that simple label unstable. Acidic luminal pH increased conductance and shifted selectivity in favor of H+, yet measured Erev values did not match the theoretical proton equilibrium voltage.
The 2026 study reframed the discrepancy. When proton currents are large, the channel can locally erode the very proton gradient used to measure it. That means Erev can move toward a proton signal and then drift back toward 0 mV as the gradient collapses.
In whole-cell recordings, acidifying the TMEM175 luminal-facing side from pH 7.4 to 4.7 increased conductance in TMEM175-expressing cells. Control cells did not show the same Erev or conductance response, making the effect channel-dependent rather than a generic acid artifact.1
Citrate Buffer Made the Proton Signal Larger
Buffer experiments were central because proton channels are difficult to measure. H+ moves quickly, buffers move more slowly, and local pH near a pore can change during the recording. Schulze et al. increased buffer capacity and compared citrate with acetate.
With 50 mM citrate, Erev peaked at +42.9 ± 6.9 mV (n = 27), more positive than the +25.6 ± 5.4 mV peak seen with 5 mM acetate (n = 18). With 50 mM acetate, the peak was smaller at +15.9 ± 9.9 mV (n = 16), consistent with acetate’s ability to contribute to pH-gradient collapse as a membrane-permeable weak acid.
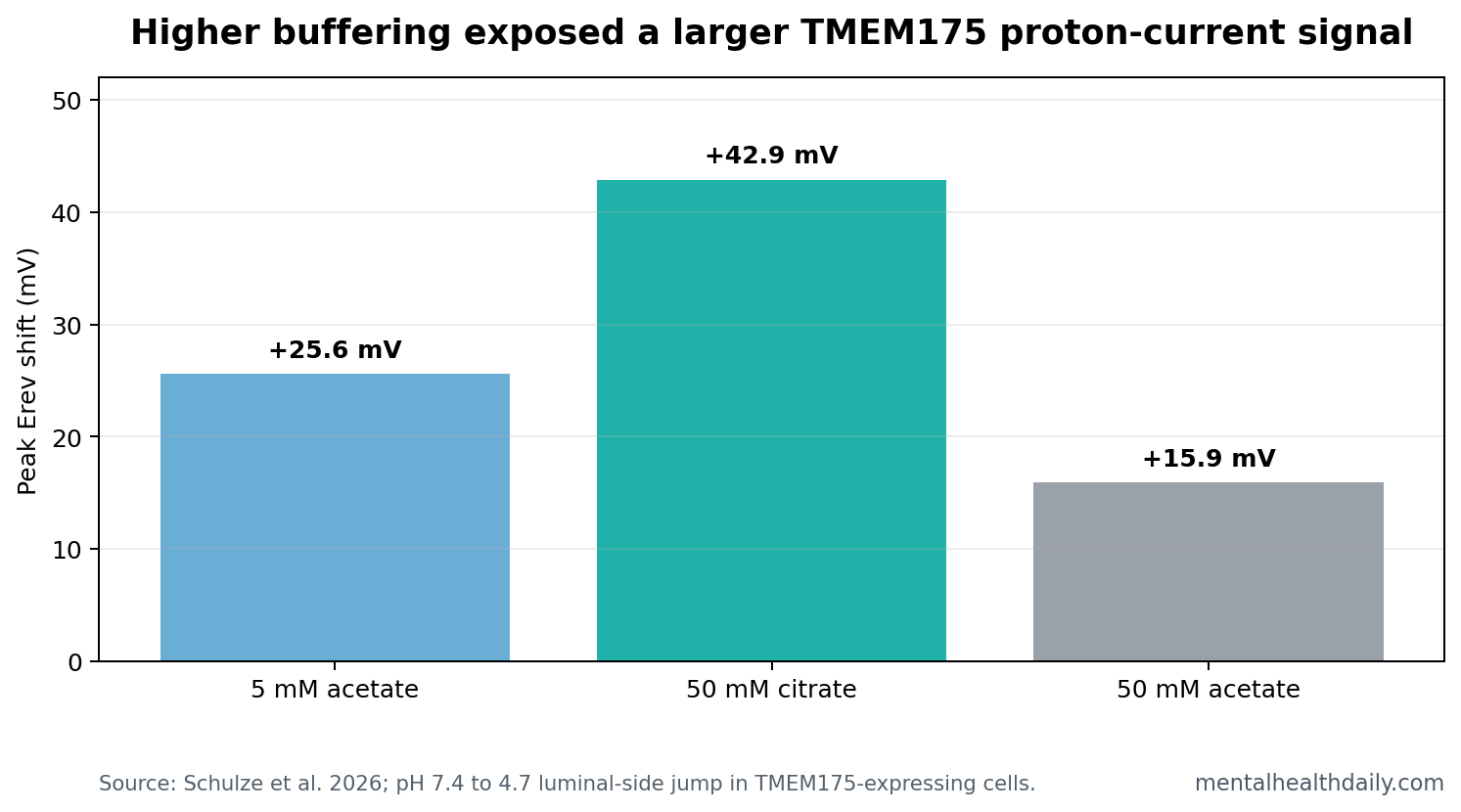
Practical interpretation: the channel’s proton behavior is real enough to appear more strongly under better proton-buffering conditions, but the measurement remains constrained by the physics of proton movement.
H57 Connected pH Sensing to Ion Flow
Molecular dynamics simulations pointed to histidine 57, or H57, on the luminal side of the open channel. H57 formed salt-bridge interactions with D279 and E282, giving the researchers a plausible structural explanation for pH-sensitive gating.
The H57Y mutant supported the structural interpretation. It reduced H+ conductance, K+ conductance, and H+/K+ selectivity in whole-cell and lysosomal electrophysiology. That matters because a disease-relevant channel mechanism is more convincing when the residue-level model changes the measured current.
Mechanism handle: TMEM175 seems to operate less like a fixed one-ion pipe and more like a pH-sensitive channel whose apparent selectivity depends on luminal acidity and local gradient collapse.
TMEM175 Connects Lysosomal pH to Parkinson’s Risk Biology
Parkinson’s disease involves dopamine loss alongside genetic, pathological, and cellular evidence for lysosomal and autophagy failure. Autophagy is the cellular cleanup process that moves damaged proteins and organelles into degradation pathways. If lysosomal pH is wrong, cleanup efficiency can fall.
TMEM175 sits in that system. Human genetic work has linked the locus to Parkinson’s disease risk, while cell and animal studies have connected TMEM175 dysfunction with lysosomal stress and α-synuclein biology.2 A pH-sensitive proton-conductance model therefore changes how the risk gene is interpreted: the problem may involve acid handling, organelle homeostasis, and potassium permeability together.
That does not make TMEM175 an immediate drug target. A channel can be disease-relevant and still hard to manipulate safely. Lysosomal pH is fundamental cell biology, so a treatment that pushes it in the wrong direction could create broad toxicity.
The Study Is Strong Mechanistic Biology, Not Patient Evidence
Evidence-strength note: this was a mechanistic electrophysiology and modeling study. It used TMEM175-expressing cells, isolated lysosomes, solid-supported membrane electrophysiology, and molecular dynamics. It did not enroll people with Parkinson’s disease or measure motor symptoms, cognition, progression, or treatment response.
The right clinical inference is restrained. TMEM175-related Parkinson’s risk should be discussed as lysosomal ion-homeostasis biology. The study does not show that changing TMEM175 activity will slow Parkinson’s disease, prevent Parkinson’s disease, or help a specific patient.
It does, however, improve the target definition. A poorly understood channel is a weak therapeutic target. A channel with pH-dependent H+ and K+ behavior, residue-level gating clues, and lysosomal electrophysiology is a clearer target for future pharmacology.
How This Changes the Search for TMEM175 Treatments
Drug discovery around TMEM175 needs to avoid a simplistic “open the channel” or “block the channel” frame. If TMEM175 helps regulate lysosomal pH, the useful intervention might depend on disease stage, variant, cell type, and baseline lysosomal acidity.
Assay design: screens should measure pH-dependent conductance across neutral and acidic conditions. A compound that looks helpful under one pH condition might behave differently in an acidic lysosome.
Variant logic: Parkinson’s-associated TMEM175 variants may need to be tested under acidic conditions, with H+ and K+ flux measured separately when possible.
Safety logic: lysosomes are everywhere. Any TMEM175 drug would need evidence that it improves vulnerable neuronal cleanup without broadly disrupting lysosomal function in other tissues.
Why Proton-Gradient Collapse Is the Key Calibration
The most technical part of the paper is also the most important for interpretation. If TMEM175 carried only K+, then acidifying the luminal side should not produce the observed transient proton-like Erev behavior. If it were a perfectly selective H+ channel under these conditions, Erev should sit much closer to the theoretical proton equilibrium voltage.
Schulze et al. argue for the middle model: acidification activates H+ conductance, but the resulting proton current rapidly erodes the local pH gradient. The measured voltage then moves back toward 0 mV because the channel changes the thing being measured.
Why that is not hand-waving: the buffer experiments support the explanation. Higher citrate buffering made the positive Erev shift larger, while acetate behaved differently because protonated acetate can cross membranes and help collapse the pH gradient. A measurement artifact became a mechanistic clue.
Why Parkinson’s researchers should care: lysosomal failure is often discussed as a broad cleanup defect. This paper narrows one piece of the cleanup system to ion flux, pH sensing, and a residue-level gate. That is the kind of mechanistic resolution needed before a risk gene becomes a credible intervention target.
What remains unresolved: the study does not tell whether Parkinson’s-associated TMEM175 variants produce too much conductance, too little conductance, mistimed conductance, or abnormal pH dependence in vulnerable neurons. Those are different therapeutic problems. A drug that corrects one variant state could worsen another if the assay is too crude.
Better translational path: variant-specific neuronal models, lysosomal pH reporters, α-synuclein clearance assays, and stress-rescue experiments should be connected before moving toward patient claims. The channel mechanism is now sharper; the disease bridge still has to be built.
That sequence would also help separate a useful biomarker from a useful drug target, since a variant can explain risk without being easy to correct pharmacologically.
A strong follow-up would test Parkinson’s-associated variants under the same pH and buffer conditions used here, then connect conductance changes to lysosomal enzyme activity, autophagy, and α-synuclein handling in neurons.
That experiment would clarify whether a variant changes the channel’s sensor, pore behavior, lysosomal set point, or stress response. Those are different drug-development problems even when they share the same gene name.
Questions About TMEM175 and Parkinson’s Disease
Is TMEM175 a potassium channel or a proton channel?
The 2026 study supports mixed behavior. TMEM175 has K+ conductance, but acidic luminal pH activates substantial H+ conductance and can collapse the proton gradient.
Does this explain Parkinson’s disease?
It explains a plausible lysosomal mechanism connected to Parkinson’s risk biology. It does not explain Parkinson’s disease as a whole.
Does this create a treatment?
No. It gives drug developers a sharper channel mechanism to test. Patient benefit would require separate disease-model and clinical evidence.
References
- Schulze T, Sprave T, Groebe C, et al. Proton-selective conductance and gating of the lysosomal cation channel TMEM175. PNAS. 2026;123:e2503909123. https://doi.org/10.1073/pnas.2503909123
- PubMed search: TMEM175 Parkinson disease lysosome alpha synuclein. https://pubmed.ncbi.nlm.nih.gov/?term=TMEM175+Parkinson+disease+lysosome+alpha+synuclein
- PubMed search: TMEM175 variants Parkinson disease risk. https://pubmed.ncbi.nlm.nih.gov/?term=TMEM175+variants+Parkinson+disease+risk
- PubMed search: lysosomal pH autophagy Parkinson disease alpha synuclein. https://pubmed.ncbi.nlm.nih.gov/?term=lysosomal+pH+autophagy+Parkinson+disease+alpha+synuclein