A 40-rat early-life visceral stress study found auditory pathway sensitization without hearing-threshold loss: neonatal colorectal distension produced IBS-like visceral hypersensitivity, normal ABR thresholds, shortened auditory brainstem latencies, increased amplitudes, and 219 differentially expressed cochlear proteins.1 The result turns IBS-like stress exposure into a gut-pain and sensory-processing question.
Research Highlights
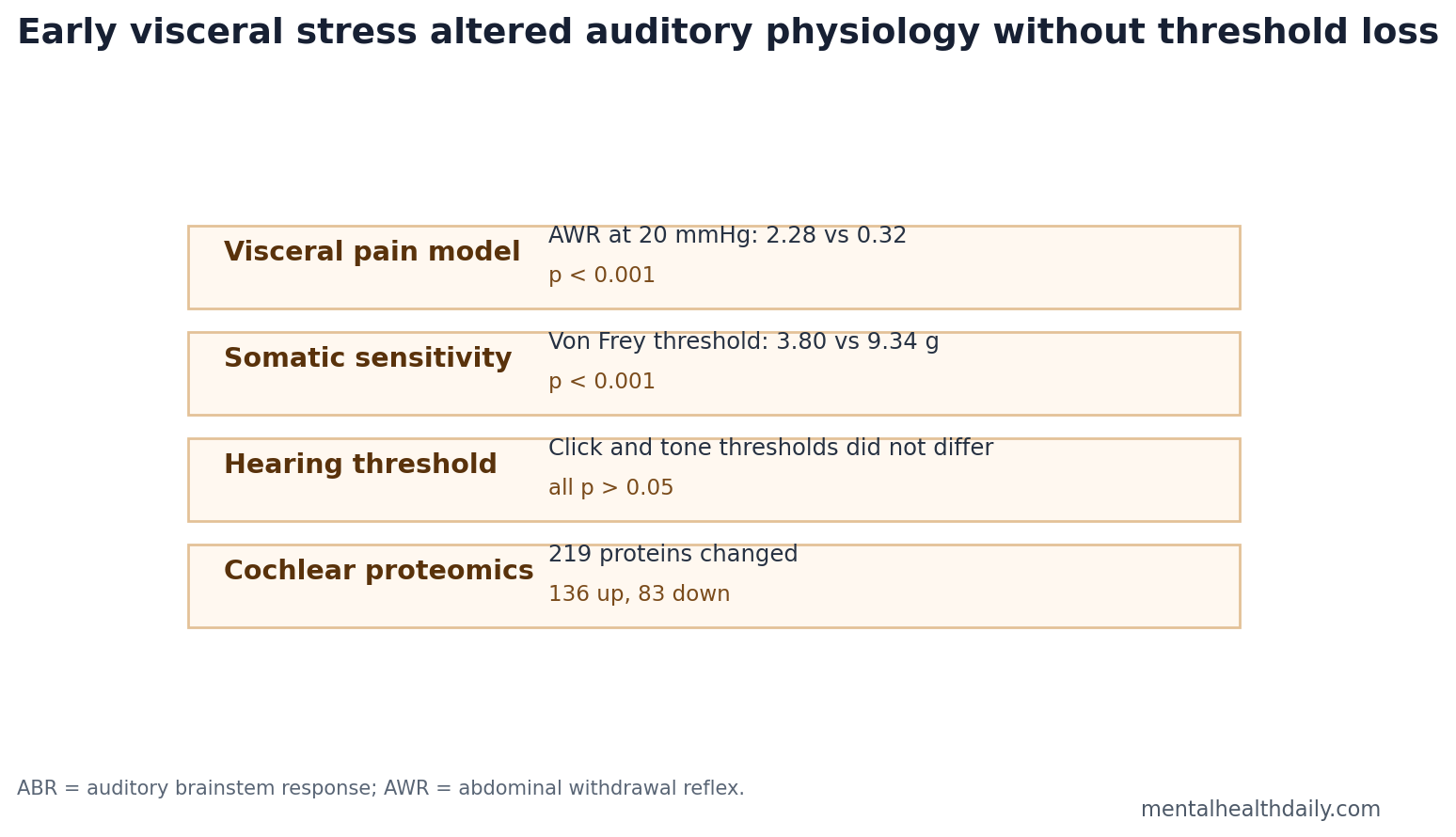
- Visceral hypersensitivity was strong: At 20 mmHg colorectal distension, IBS-model rats had AWR scores of 2.28 vs. 0.32 in controls; at 80 mmHg, scores were 3.60 vs. 2.12, both p < 0.001.1
- Somatic sensitivity also increased: Hindpaw mechanical withdrawal thresholds were lower in IBS-model rats, 3.80 vs. 9.34 g, p < 0.001.1
- Hearing thresholds stayed normal: ABR thresholds did not differ for click or 4, 8, 16, and 24 kHz tone bursts, all p > 0.05.1
- Brainstem responses looked sensitized: IBS-model rats showed shorter ABR latencies and larger wave amplitudes than controls across most waves and frequencies.1
- Cochlear proteins shifted: Proteomics found 219 differentially expressed proteins, including 136 upregulated and 83 downregulated proteins, enriched in vesicle, synaptic, mitochondrial, and inflammatory pathways.1
Auditory brainstem response (ABR) is an electrical recording that tracks how sound signals move from the auditory nerve through brainstem relays. A normal threshold with shorter latencies and larger amplitudes points toward stronger or faster pathway responsiveness rather than better hearing.
Early-life visceral stress in this study meant repeated neonatal colorectal distension from postnatal day 8 to 21. That procedure is a standard rat model for IBS-like visceral hypersensitivity, not a model of psychological trauma or human childhood adversity in full.2
Neonatal Colorectal Distension Produced IBS-Like Hypersensitivity
Zhang et al. assigned male Sprague-Dawley rats to an IBS-model group, n = 20, or control group, n = 20. At 7 weeks, the IBS-model group weighed less than controls, 221.78 vs. 282.50 g, p < 0.001, and showed clear visceral and somatic hypersensitivity.1
- 20 mmHg distension: AWR score 2.28 vs. 0.32, p < 0.001.
- 40 mmHg distension: AWR score 3.10 vs. 0.93, p < 0.001.
- 60 mmHg distension: AWR score 3.40 vs. 1.77, p < 0.001.
- 80 mmHg distension: AWR score 3.60 vs. 2.12, p < 0.001.
- Von Frey threshold: 3.80 vs. 9.34 g, p < 0.001.
Abdominal withdrawal reflex (AWR) is a graded behavioral score for abdominal contraction and body response during colorectal distension. The pattern confirmed that the manipulation did what it was supposed to do: create persistent visceral hypersensitivity.
The model is worth separating from broad “early adversity” language. Neonatal colorectal distension is a targeted gut-pain stressor, not a social-defeat, maternal-separation, or fear-conditioning protocol. It mainly tests whether repeated visceral input during a sensitive developmental window can leave the adult animal with a lower threshold for gut and body-pain signaling.
That narrower design is also the reason the auditory result is interesting. If the manipulation had produced only abdominal sensitivity, the study would have repeated the standard IBS-model finding. Instead, the same animals showed a second sensory-system phenotype: normal detection thresholds but altered neural response timing after sound reached the auditory pathway.
Auditory Thresholds Were Normal, but ABR Timing and Amplitude Changed
The important calibration is that hearing threshold was not the finding. ABR thresholds did not differ between IBS-model rats and controls for click or tone-burst stimuli at 4, 8, 16, and 24 kHz. Click thresholds were 22.00 vs. 21.50 dB, p = 0.736; all frequency comparisons were nonsignificant.1
Latency and amplitude moved instead. IBS-model rats showed shorter latencies across many waves and frequencies, especially early waves and higher frequencies, plus larger ABR amplitudes across most waves and frequencies. That combination points to auditory pathway hyperexcitability rather than peripheral hearing loss.
Latency is the time between the sound stimulus and a measured electrical wave. Shorter latency can mean faster neural conduction, altered synaptic timing, or stronger synchronized firing along the auditory nerve and brainstem pathway. Amplitude is the size of the response. Larger amplitude can mean that more neural elements are firing together or that the system is more excitable at the tested sound level.
The rat data therefore do not say “IBS improves hearing.” They say the threshold for detecting sound stayed about the same while the downstream response looked amplified and faster. That distinction matters because sensory complaints in chronic-pain conditions often involve gain control, salience, and central processing rather than a simple damaged-ear model.
Central gain is the practical concept: a sensory pathway can turn up response strength after repeated stress or pain input even when the peripheral organ still detects stimuli at ordinary thresholds. In this study, normal thresholds argue against a simple cochlear-damage explanation, while faster and larger ABR waves keep the focus on response regulation along the auditory pathway.
219 Cochlear Proteins Shifted After Early Visceral Stress
The proteomic analysis identified 4,417 quantified proteins in inner-ear tissue. Using fold-change thresholds above 1.3 or below 0.77 plus p < 0.05, the researchers found 219 differentially expressed proteins: 136 upregulated and 83 downregulated.1
Enrichment analyses pointed toward vesicle-mediated transport, cytokine-mediated signaling, synaptic vesicles, mitochondrial and Golgi membranes, SNARE and GTPase binding, and inflammatory signaling pathways. KEGG pathway results included MAPK, PI3K-Akt, TNF, HIF-1, IL-17, Ras signaling, apoptosis, and endocytosis.1
Mechanistic read: the inner ear may not be a passive downstream target. Inflammatory, synaptic, vesicle-trafficking, and mitochondrial changes could plausibly alter cochlear or auditory-nerve responsiveness after early visceral stress.
Several pathway labels point in the same general direction without proving a single causal chain:
- Cytokine and TNF signaling: immune communication pathways that can change support-cell and neural excitability in stressed tissue.
- Synaptic vesicle and SNARE terms: machinery for neurotransmitter release, which fits an altered neural-response phenotype better than a pure structural-hearing-loss story.
- Mitochondrial pathways: energy-handling systems that can influence how sensory cells and neurons tolerate repeated activation.
- Endocytosis and vesicle transport: cellular trafficking processes that can change receptor recycling, membrane signaling, and inflammatory responses.
Those categories are hypothesis-generating. Proteomics measured abundance patterns in cochlear tissue after the phenotype was already present; it did not block TNF, rescue mitochondrial function, or isolate a single auditory-cell type. The strongest claim is a coherent tissue-level signal, not a proved drug target.
Human IBS Studies Already Hint at Auditory Processing Changes
Clinical IBS is often framed around abdominal pain, bowel habits, anxiety, depression, sleep disturbance, and chronic-pain comorbidity. Smaller human studies have also reported altered auditory event-related potentials, including increased N100 and P1 amplitudes in IBS patients.3,4
Zhang et al. did not test human IBS patients. The value of the rat study is mechanistic: it shows that a gut-stress model can produce auditory brainstem changes and cochlear molecular changes in the same animals.
Event-related potentials are scalp-recorded brain responses time-locked to a stimulus. N100 and P1 are early auditory-processing components, not questionnaires about annoyance or sound tolerance. The human ERP findings and the rat ABR findings therefore occupy neighboring levels of the same problem: objective auditory-system responses can differ in IBS-relevant states even when the main clinical diagnosis is gastrointestinal.
The translational bridge is still incomplete. Human IBS studies would need to show whether altered auditory responses track visceral pain severity, widespread pain sensitivity, anxiety, sleep disturbance, inflammatory markers, or medication exposure. Without that mapping, it would be easy to overread a cross-modal sensory signal as a direct ear disease.
What This Rat Study Can and Cannot Support
Supported: neonatal visceral stress in male rats produced persistent visceral and somatic hypersensitivity, normal auditory thresholds, altered ABR latency/amplitude, and cochlear proteomic changes.
Not supported: a human claim that IBS causes hearing loss, tinnitus, misophonia, or auditory hypersensitivity. ABR testing lacked anatomical resolution, proteomics did not prove causality, and recordings were taken at a single time point.
Best next test: human IBS studies should pair auditory psychophysics, ABR or ERP measures, pain-sensitization phenotyping, inflammatory markers, and symptom severity to see whether a gut-inner-ear signal exists outside animal models.
Sex and development checks: this study used male rats, so it cannot tell whether the same pathway appears in females, even though IBS is clinically common in women. Repeating the work with female rats, multiple postnatal windows, and later adult time points would help separate a durable sensory-development effect from a narrower male-model finding.
Another useful next step would be intervention testing. If anti-inflammatory, neuromodulatory, microbiome, or stress-buffering interventions reduce visceral hypersensitivity but leave ABR changes intact, the gut and auditory effects may diverge. If both move together, the case for a shared sensitization pathway becomes stronger.
The near-term reader takeaway is narrow but real: early gut-pain stress can leave measurable signatures outside the gut in an animal model. That supports broader sensory-system testing in IBS research, while keeping human claims about hearing symptoms, tinnitus, or sound intolerance off the table until human data directly measure them.
Questions About IBS and Auditory Sensitivity
Did the rats lose hearing?
No. Thresholds were comparable. The signal was faster and larger ABR responses, which points to excitability or sensitization rather than threshold loss.
Is this the same as human sound sensitivity?
No. ABR changes are physiological signals. Human sound sensitivity involves perception, attention, distress, and context, which this rat model did not measure.
Why mention the cochlea in a stress article?
The cochlea contains immune and support-cell systems that can respond to systemic inflammation and metabolic stress. This study suggests early visceral stress may reach auditory biology through those pathways.
References
- Zhang LY, Li XL, Jing YY, Yu LS. Early life visceral stress induces auditory brainstem sensitization and cochlear proteomic alterations in a rat model of irritable bowel syndrome. Neurobiology of Stress. 2026;42:100812. doi:10.1016/j.ynstr.2026.100812
- Al-Chaer ED, Kawasaki M, Pasricha PJ. A new model of chronic visceral hypersensitivity in adult rats induced by colon irritation during postnatal development. PubMed
- Blomhoff S, et al. Auditory event-related potentials in irritable bowel syndrome. PubMed
- Berman SM, et al. Cerebral response to auditory stimulation in irritable bowel syndrome. PubMed
- Camilleri M. Diagnosis and treatment of irritable bowel syndrome. PubMed