
A 2026 iScience study recorded 27 substantia nigra neurons during deep-brain-stimulation surgery in 11 Parkinson’s disease patients and found that prior reward shaped both putative dopamine-neuron firing and the next reaction time: responses were faster after a +$10 outcome than after neutral or negative outcomes (0.916 s vs. 1.017 s, p = 0.0175).1
Research Highlights
- Direct human recordings were rare: researchers recorded 27 substantia nigra neurons across 13 sessions in 11 Parkinson’s disease patients undergoing DBS surgery.1
- Most neurons were putative dopamine cells: 22 of 27 neurons were classified as putative dopamine neurons based on firing rate below 15 Hz and waveform width above 0.8 ms.1
- Prior reward changed expectation firing: previous reward outcome predicted firing rate during reward expectation (t(2217) = 5.412, p = 6.91e-8), while previous action and previous reward-prediction error did not.1
- 8 of 22 neurons carried the single-cell signal: individual-neuron models found a significant previous-outcome effect in 8 putative dopamine neurons.1
- Behavior matched the neural direction: median reaction time was faster after reward than after neutral or negative outcomes (0.916 s vs. 1.017 s, p = 0.0175).1
Substantia nigra is a midbrain region rich in dopamine neurons, especially relevant to movement, learning, reward, and Parkinson’s disease. Dopamine helps the brain update expectations, energize action, and learn from outcomes; the “pleasure chemical” shortcut misses most of the biology.
Reward prediction error means the gap between expected and actual reward. Classic dopamine work showed that dopamine-neuron firing can shift from unexpected reward to cues that predict reward.2 Imtiaz et al. asked a narrower human question: does the immediately prior outcome shape substantia nigra firing before the next decision?
That narrower question is clinically interesting because many psychiatric and neurological symptoms involve what happens after recent success or failure. A reward can energize the next action; a loss can slow responding, promote switching, or change expected value. The study tested that short-horizon carryover directly in human neurons rather than inferring it only from behavior or brain imaging.
27 Neurons Were Recorded During Parkinson’s DBS Surgery
The study used intraoperative microelectrode recording during deep-brain-stimulation implantation. Deep brain stimulation (DBS) is a neurosurgical treatment in which electrodes are implanted to modulate abnormal circuit activity, commonly in movement disorders such as Parkinson’s disease.
Researchers recorded 27 neurons across 13 sessions in 11 patients. Of those, 22 were classified as putative dopamine neurons and 5 as presumed GABAergic interneurons. The classification used firing rate and waveform width because direct neurochemical labeling is not possible during this kind of human surgery.1
The putative-dopamine label is important. A neuron with low firing rate and wide waveform is more likely to be dopaminergic in this setting, but the researchers could not stain the cell or measure dopamine release from the same unit. The study therefore supports human substantia nigra reward-history coding; it should not be read as cell-by-cell chemical proof.
Prior +$10 Outcomes Increased Expectation-Period Firing
Patients played a two-armed bandit task involving reward, punishment, and mixed blocks. Choices were influenced strongly by the immediately previous outcome, fitting a win-stay/lose-switch style pattern rather than random choice.1
The neural result was strongest before feedback. Previous reward outcome predicted expectation-period firing in putative dopamine neurons (t(2217) = 5.412, p = 6.91e-8). Previous motor action did not (p = 0.8901), and previous reward-prediction error did not (p = 0.3264). At the single-neuron level, 8 of 22 putative dopamine neurons showed a significant previous-outcome effect.
That model separates reward history from simple movement repetition. If firing only reflected which button the patient had pressed, previous action should have carried the effect. It did not. The stronger signal was the prior outcome itself, which makes the result more relevant to expectation and motivation than to motor habit.
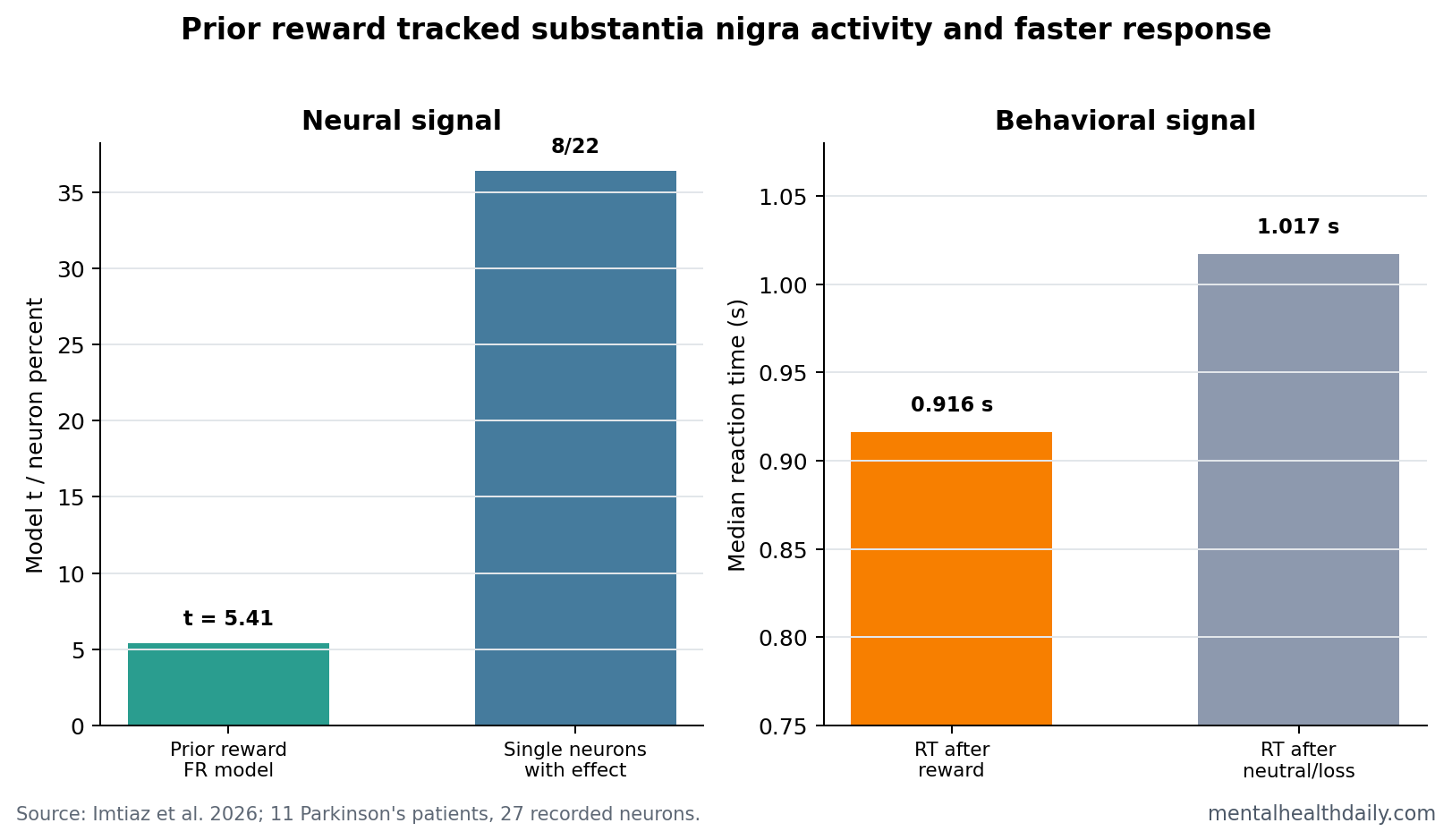
Reaction Time Was Faster After Reward
The behavioral result moved in the same direction as the neural result. Median reaction time was 0.916 seconds after +$10 outcomes compared with 1.017 seconds after neutral or negative outcomes (p = 0.0175).1
Motivational vigor means how energetically and quickly an organism acts when reward seems available. Niv et al. modeled tonic dopamine as a control signal for vigor: higher expected reward should justify faster responding because time spent waiting has an opportunity cost.3 The 2026 human recording study fits that framework without proving the whole model.
The size of the reaction-time difference was modest but concrete: about 101 milliseconds faster after reward. In a lab task, 101 milliseconds is enough to show that recent outcome changed readiness to act. In clinical life, the same principle could scale into larger patterns such as apathy, reward sensitivity, medication-state effects, or addiction-related cue responsiveness, but those translations need separate evidence.
The Study Found More Reward-History Encoding Than Feedback Encoding
Post-feedback reward encoding was weaker and more heterogeneous. Seven of 22 putative dopamine neurons showed some feedback modulation by uncorrected tests, but only 1 of 22 survived more stringent cluster-based permutation testing. Some neurons increased firing after reward, while others decreased.1
The strongest group signal was not “dopamine fired after reward.” Recent reward history shaped sustained firing during the next expectation period. In normal decision language, the last outcome changed the baseline from which the next possible outcome would be evaluated.
That distinction also fits the behavioral model. Patients were not choosing randomly; their choices were captured by reward-history strategies such as win-stay/lose-switch. Neuronal activity and choice behavior therefore pointed in the same direction: the immediately preceding outcome carried forward into the next decision.
Parkinson’s Surgery Limits Generalization
Evidence-strength note: this was a small intraoperative study in Parkinson’s disease patients, not a healthy-volunteer experiment. Chronic dopaminergic degeneration, surgical context, medication history, small neuron counts, and brief task duration all limit generalization.
The study still matters because direct human single-unit dopamine recordings are scarce. Zaghloul et al. previously showed human substantia nigra neurons responding to unexpected financial rewards.4 Imtiaz et al. add a reward-history signal that looks closer to expectation and response vigor than simple feedback detection.
Parkinson’s disease also cuts in 2 directions. It limits generalization because dopamine neurons are affected by the disease process. It also makes the sample clinically relevant because motivation, action initiation, and reward learning are core problems in Parkinson’s disease and in dopaminergic treatment states.
Motivation Disorders Need More Than a Dopamine Slogan
The clinical translation should stay narrow. Depression, addiction, apathy, psychosis, Parkinson’s disease, and compulsive behavior all involve motivation and reward in different ways. A 101-millisecond reaction-time shift after a +$10 trial inside a DBS task does not diagnose those conditions.
What it does show is a plausible micro-scale mechanism: recent reward can tune human substantia nigra activity before the next choice, and that tuning tracks faster action. That is the kind of signal that could matter when reward history, effort, and motivation become clinically disrupted.
For depression and addiction research, the useful bridge is not that these 11 patients explain those disorders. It is that reward history can be represented in human midbrain firing before action. Future studies can ask whether that signal is blunted in apathy, exaggerated by stimulant or dopaminergic medication states, or altered when rewards become compulsive rather than adaptive.
The task design also separates immediate reward history from long-run personality or motivation traits. Each trial created a short memory of reward, nonreward, or punishment, then measured how the next decision unfolded. That trial-by-trial structure is why the result speaks to rapid updating, not a stable “motivated person” trait.
Clinically, that distinction helps avoid dopamine overstatement. A patient with low motivation may have impaired reward learning, low expected value, motor slowing, medication effects, depression, sleep disruption, or environmental reward scarcity. Substantia nigra reward-history firing is one possible mechanism inside that larger system.
For Parkinson’s disease specifically, the result also sits beside medication timing and DBS state. Dopaminergic drugs can change reward learning and impulse-control risk, while DBS surgery targets motor circuits. A human single-unit reward-history signal gives researchers a finer tool for asking how those treatments affect motivation.
The next useful experiment would vary medication state, reward size, and effort cost while recording the same expectation-period signal.
A clinically stronger design would also separate motor speed from motivation. Parkinson’s patients can respond slowly because movement execution is impaired, because expected reward is weak, or because the medication state changes both at once. Recording neurons while effort cost and motor demand are controlled would make the motivational interpretation cleaner.
Questions About Dopamine Reward-History Signals
Did the study prove dopamine causes motivation?
No. It found an association between prior reward, putative dopamine-neuron firing, and faster reaction time in a small surgical sample. Causality and clinical translation require other designs.
Why were Parkinson’s patients studied?
Direct human substantia nigra recording is usually possible only during neurosurgery. Parkinson’s DBS surgery creates a rare ethical and technical window for single-neuron recording.
Was previous action the signal?
No. Previous motor action did not predict expectation-period firing in the final model. Previous reward outcome did.
References
- Imtiaz Z, et al. Sustained activity of human substantia nigra neurons reflect prior rewards. iScience. 2026. https://doi.org/10.1016/j.isci.2026.115458
- Schultz W, Dayan P, Montague PR. A neural substrate of prediction and reward. Science. 1997. https://doi.org/10.1126/science.275.5306.1593
- Niv Y, et al. Tonic dopamine: opportunity costs and the control of response vigor. Psychopharmacology. 2007. https://doi.org/10.1007/s00213-006-0502-4
- Zaghloul KA, et al. Human substantia nigra neurons encode unexpected financial rewards. Science. 2009. https://doi.org/10.1126/science.1167342