
A 2026 whole-genome sequencing preprint found rare variant signals in 2 multiplex obsessive-compulsive disorder (OCD) families: an NPY5R missense variant in 10 of 15 affected people in 1 pedigree, plus DLGAP1 and MAPK8IP3 signals that tracked all 3 sequenced cases in a branch of a second pedigree.1 The finding is a candidate-gene result, not a clinical genetic test.
Research Highlights
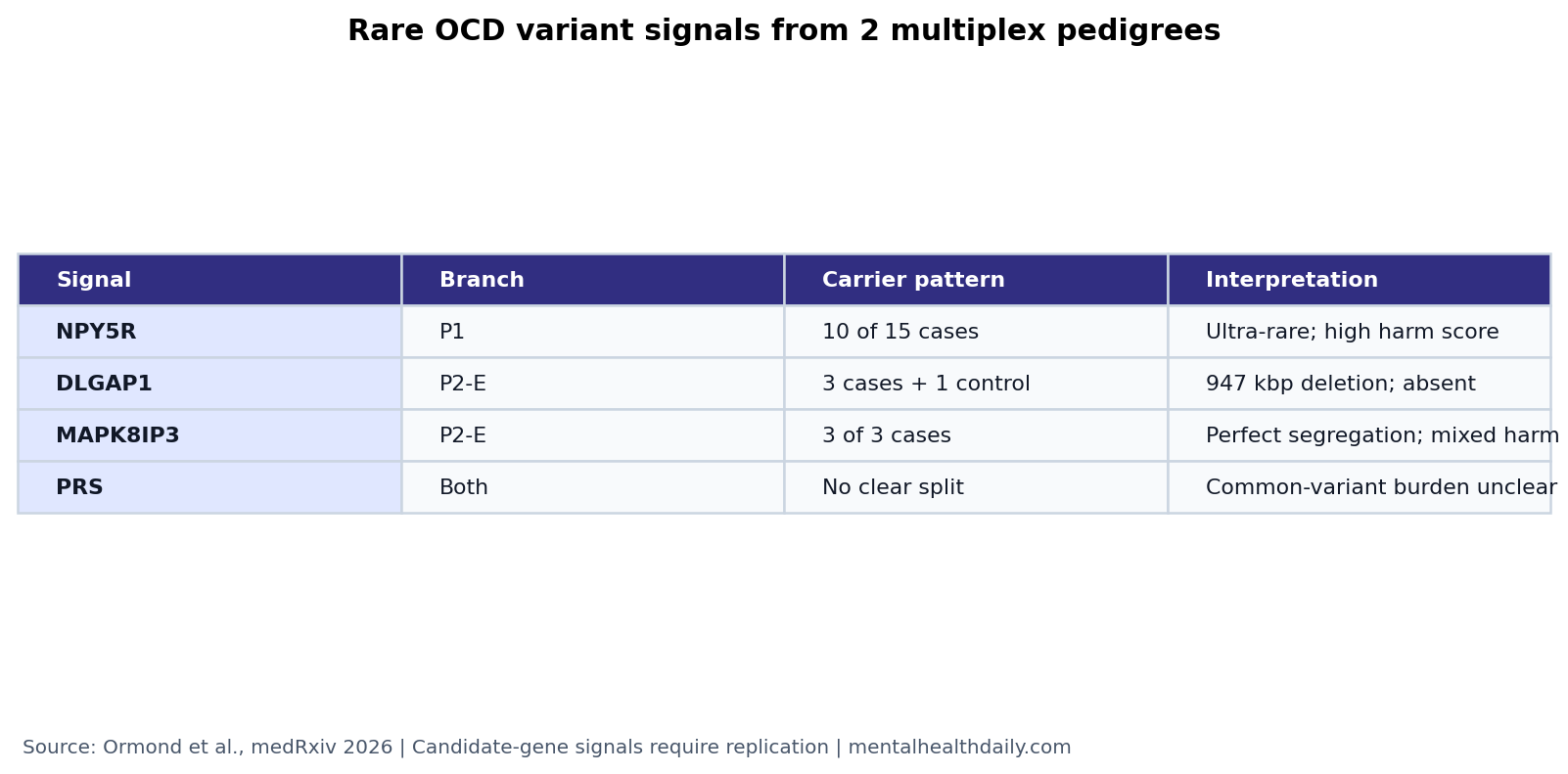
- Family sequencing found 3 main signals: Ormond et al. sequenced 25 people across 2 OCD pedigrees and prioritized rare variants in NPY5R, DLGAP1, and MAPK8IP3.1
- NPY5R was the strongest P1 candidate: an ultra-rare, highly deleterious NPY5R missense variant was carried by 10 of 15 OCD cases in the first pedigree.1
- DLGAP1 and MAPK8IP3 tracked a P2 branch: the Founder E branch had a 947 kbp DLGAP1 deletion in all 3 cases and 1 control, while MAPK8IP3 was carried by all 3 cases.1
- Common-variant burden did not explain the split: polygenic risk scores for OCD, Tourette syndrome, and panic disorder showed no clear carrier vs. noncarrier pattern in the families.1
- Replication is the hard gate: the study is a preprint and the strongest claims come from small family branches, so independent OCD cohorts still have to test whether these genes carry broader risk.2
Obsessive-compulsive disorder is a psychiatric condition defined by intrusive obsessions, compulsions, or both. Twin and family studies support substantial inherited liability, but OCD genetics has not behaved like Huntington’s disease or another single-gene disorder. Most risk appears distributed across many common variants, rare coding variants, rare copy number variants, and family-specific genetic background.
The Ormond preprint sits in the family-based rare-variant lane. That lane can find strong local signals in densely affected families, but it has a narrow burden of proof: a variant can track disease inside a family and still fail to generalize to OCD more broadly.
25 People Across 2 OCD Pedigrees Were Sequenced
Ormond et al. used whole-genome sequencing, which reads genetic variation across the genome rather than only selected exons or common genotyping markers. The study included 15 sequenced people from pedigree P1 and 10 from pedigree P2, with OCD diagnoses assigned through structured clinical assessments and best-estimate consensus methods.1
The analysis looked for 3 kinds of genetic signal:
- Single-nucleotide variants and indels: small DNA changes that can alter a protein, disrupt a splice site, or change gene function.
- Copy number variants: larger deletions or duplications of DNA segments that can remove or duplicate gene material.
- Polygenic risk scores: genome-wide scores built from many common variants, used here to ask whether family members without the rare candidate variant had higher common-variant liability.
The key prioritization method was BICEP, a Bayesian rare-variant scoring framework that combines predicted variant harm with co-segregation inside a pedigree. In plain English, it asks whether a rare variant is both biologically suspicious and inherited with the diagnosis more often than expected.
NPY5R Was Carried by 10 of 15 OCD Cases in Pedigree P1
In pedigree P1, the strongest candidate was an ultra-rare, highly deleterious missense variant in NPY5R, a gene for neuropeptide Y receptor 5. A missense variant changes one amino acid in a protein; it is not automatically damaging, but prediction tools can flag variants that are unusually likely to alter protein function.
The carrier pattern was substantial but incomplete: 10 of 15 affected people carried the NPY5R variant, while 5 affected people did not.1 That makes the signal biologically interesting, not decisive. If the variant contributes to risk in this family, it is unlikely to be the only route into OCD for the pedigree.
NPY5R is plausible because neuropeptide Y signaling is involved in stress, anxiety, feeding, and emotional regulation. Prior association work has linked NPY receptor variation to panic disorder and substance-related phenotypes, though larger OCD genome-wide studies have not established NPY5R as a confirmed OCD locus.3,4
Ormond et al. also reran BICEP using any anxiety disorder as the phenotype because several family members had anxiety-related diagnoses. The NPY5R variant did not co-segregate better with anxiety than with OCD, which keeps the candidate aligned with OCD in this family rather than a broader anxiety-only explanation.1
DLGAP1 and MAPK8IP3 Co-Segregated in One P2 Branch
Pedigree P2 had 2 major founder branches, so the researchers analyzed the branches separately. The most interesting pattern came from the Founder E branch, where 2 candidate signals emerged.
DLGAP1 encodes a postsynaptic density scaffold protein. Postsynaptic density means the dense protein machinery on the receiving side of a synapse, where receptors and signaling proteins organize communication between neurons.
In P2’s Founder E branch, researchers found a 947 kbp deletion overlapping the first 3 exons in the untranslated region of DLGAP1. It appeared in all 3 cases and 1 control, was absent from gnomAD, and had a CADD-SV Phred score of 12.97, meaning it ranked in roughly the top 5% of predicted deleterious structural variants.1
The control carrier is important. It weakens any simple one-variant-one-diagnosis claim, while still allowing a risk-modifier interpretation. In psychiatric genetics, incomplete penetrance is ordinary: a risk variant can increase probability without guaranteeing disease.
MAPK8IP3 encodes JIP3, a neuronal protein involved in axonal transport and neurodevelopment. Axonal transport is the movement of cargo along nerve-cell projections, which helps neurons maintain long-range communication. In the same Founder E branch, an ultra-rare MAPK8IP3 missense variant was carried by all 3 cases and achieved the maximum possible co-segregation score, though predicted deleteriousness evidence was mixed.1
OCD Genetics Already Points Beyond Common Variants Alone
Large common-variant studies are finally gaining traction in OCD. Strom et al. reported 30 genome-wide loci associated with OCD in 2025, showing that common variants can identify broad biological architecture when sample sizes are large enough.2
Rare-variant studies answer a different question. Halvorsen et al. reported a burden of rare damaging coding variants in OCD, and later work found a burden of rare copy number variants, including enrichment in deletions affecting loss-of-function-intolerant genes.5,6 Those findings make the DLGAP1 deletion more plausible because DLGAP1 has a pLI score of 1.00, a common measure indicating strong intolerance to loss-of-function variation.
De novo coding studies also suggest overlap between OCD, Tourette disorder, and autism biology. Cappi et al. found damaging coding mutations associated with OCD and overlapping with Tourette disorder and autism, while Gazzellone et al. previously reported pediatric OCD copy number variant evidence involving DLGAP1.7,8
The Ormond study therefore does not stand alone. Its genes are not random names pulled from a pedigree scan; they sit near prior OCD, tic-disorder, autism, synaptic, and neurodevelopmental evidence. The correct conclusion is still narrower than “OCD genes found.”
Polygenic Risk Scores Did Not Rescue the Unexplained Cases
One obvious question is whether affected noncarriers in P1 had higher ordinary common-variant risk. Ormond et al. tested that with polygenic risk scores for OCD, Tourette syndrome, and panic disorder, selected because those diagnoses or related traits appeared in the family context.1
The answer was no clear pattern. In P1, the 5 affected noncarriers of the NPY5R variant did not show noticeably different polygenic risk profiles from carriers. Across both pedigrees, family members were spread reasonably across the background distribution.
That null does not prove common variants are irrelevant. Polygenic risk scores are noisy, ancestry-sensitive, and built from GWAS that still capture only part of OCD liability. It does mean this particular dataset did not show a clean “rare variant carriers vs. common-variant burden” split.
What This Preprint Can and Cannot Support
Evidence-strength note: this is a medRxiv preprint involving 25 sequenced people across 2 families. It can prioritize candidate genes for replication and functional follow-up. It cannot establish clinical genetic testing, individual risk prediction, or causality for OCD.
Three caveats shape the interpretation:
- Family-level co-segregation is not population association. A variant can matter in 1 family because of founder effects, background genetics, or chance.
- Incomplete carrier patterns are built into the result. NPY5R missed 5 of 15 affected people in P1; DLGAP1 included 1 control carrier in P2.
- Mechanism remains inferential. NPY5R suggests stress and energy-homeostasis biology, while DLGAP1 and MAPK8IP3 suggest synaptic scaffolding and neurodevelopmental transport. Functional assays still have to test those mechanisms directly.
The most useful read is that OCD genetics is becoming less abstract. Common-variant GWAS names broad loci, rare-variant burden studies show that damaging coding and structural variants matter, and family sequencing can point to specific genes worth testing. NPY5R, DLGAP1, and MAPK8IP3 are candidates in that pipeline, not settled answers.
Questions About Rare OCD Variant Findings
Does this mean NPY5R, DLGAP1, or MAPK8IP3 cause OCD?
No. The study found family-level candidate signals. Causality would require replication in independent OCD datasets and functional work showing how the variants alter relevant biology.
Could someone get tested for these variants to estimate OCD risk?
No. The evidence is not ready for clinical genetic testing. The paper itself is a preprint, the sample is small, and the variants are not established diagnostic markers.
Why study families instead of only large cohorts?
Rare high-impact variants may be too uncommon to detect efficiently in ordinary case-control studies. Multiplex families can expose variants that track disease locally, then larger studies can test whether those genes matter more broadly.
What would make the evidence stronger?
Independent OCD cohorts showing excess damaging variation in the same genes, functional assays showing altered NPY5R, DLGAP1, or MAPK8IP3 biology, and animal or cellular models linking those changes to compulsive or anxiety-related phenotypes would all strengthen the claim.
References
- Rare protein-disrupting variants in NPY5R, DLGAP1 and MAPK8IP3 segregate with OCD in two multiplex pedigrees. Ormond C, Cap M, Chang YC, et al. medRxiv. 2026. doi:10.64898/2026.04.21.26350600
- Genome-wide analyses identify 30 loci associated with obsessive-compulsive disorder. Strom NI, Gerring ZF, Galimberti M, et al. Nature Genetics. 2025;57(6):1389–1401. doi:10.1038/s41588-025-02189-z
- Chromosome 4q31-34 panic disorder risk locus: association of neuropeptide Y Y5 receptor variants. Domschke K, Hohoff C, Jacob C, et al. American Journal of Medical Genetics Part B. 2008;147B(4):510–516. doi:10.1002/ajmg.b.30629
- The NPY system in stress, anxiety and depression. Heilig M. Neuropeptides. 2004;38(4):213–224. doi:10.1016/j.npep.2004.05.002
- Exome sequencing in obsessive-compulsive disorder reveals a burden of rare damaging coding variants. Halvorsen M, Samuels J, Wang Y, et al. Nature Neuroscience. 2021;24(8):1071–1076. doi:10.1038/s41593-021-00876-8
- A burden of rare copy number variants in obsessive-compulsive disorder. Halvorsen MW, de Schipper E, Backman J, et al. Molecular Psychiatry. 2025;30(4):1510–1517. doi:10.1038/s41380-024-02763-7
- De novo damaging DNA coding mutations are associated with obsessive-compulsive disorder and overlap with Tourette’s disorder and autism. Cappi C, Oliphant ME, Peter Z, et al. Biological Psychiatry. 2020;87(12):1035–1044. doi:10.1016/j.biopsych.2019.09.029
- Uncovering obsessive-compulsive disorder risk genes in a pediatric cohort by high-resolution analysis of copy number variation. Gazzellone MJ, Zarrei M, Burton CL, et al. Journal of Neurodevelopmental Disorders. 2016;8:36. doi:10.1186/s11689-016-9170-9