
A 2026 PET-fMRI study of 20 healthy adults found that GABAA receptor binding tracked speech-production brain activity across frontal, temporal, parietal, putaminal, supplementary motor, and cerebellar regions, with peak GABA-BOLD correlations reaching Rs = 0.9.1 Human speech control looks less like a pure motor-output problem and more like a task-specific balance between inhibition, dopamine, and left-striatal circuit tuning.
Research Highlights
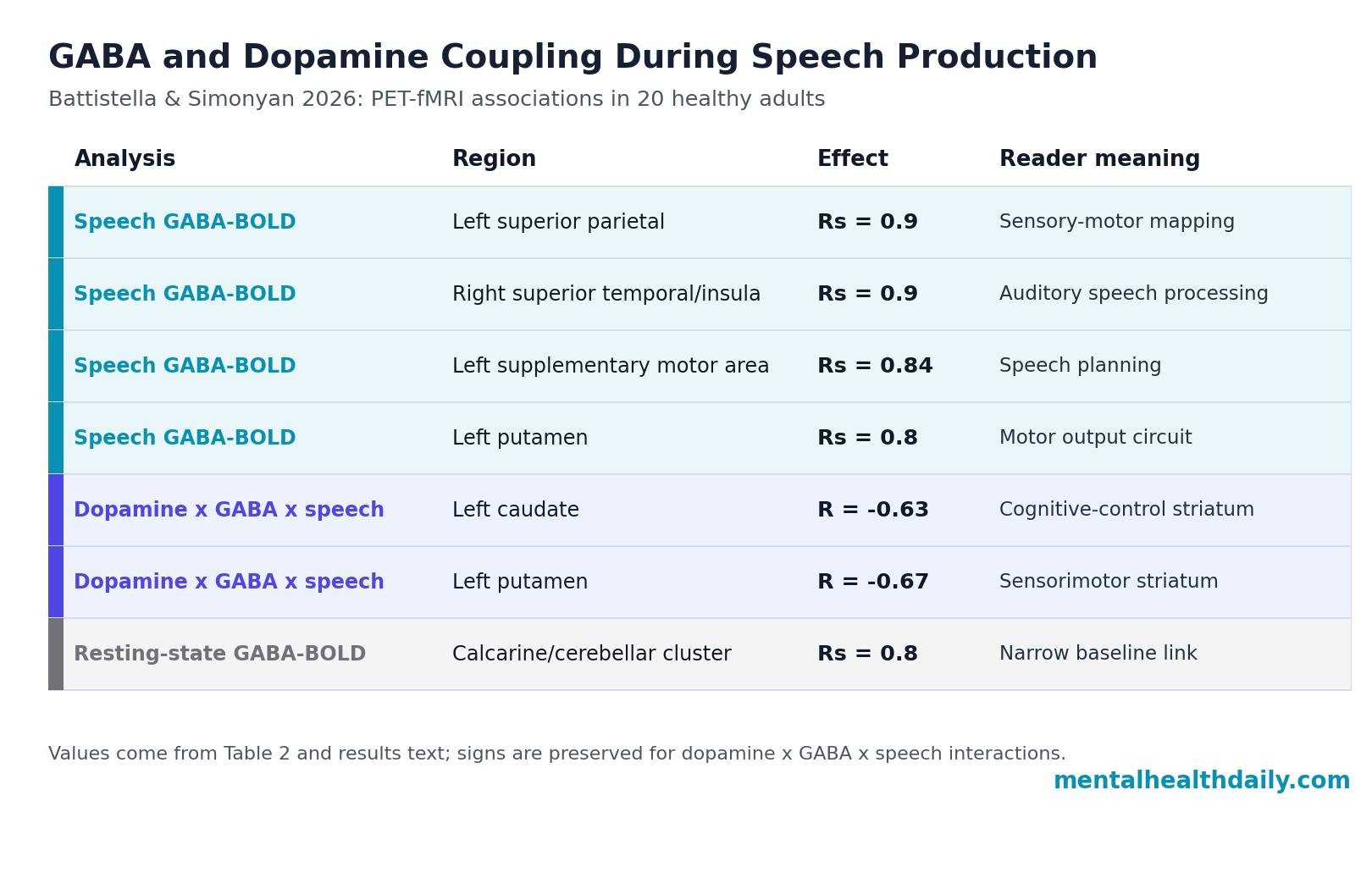
- Speech activity tracked GABA strongly: GABAA receptor binding correlated with speech-production BOLD activity in core speech-control regions, including peak Rs = 0.9 in the left superior parietal lobule and right superior temporal/insula cluster.1
- The left SMA signal was especially tight: The left supplementary motor area showed Rs = 0.84, p = 2 x 10−5, linking inhibitory receptor binding to a region used for speech planning and motor sequencing.1
- Dopamine entered the model in the striatum: Speech-related activity was modulated by GABA-dopamine interaction in the left caudate (R = −0.63, p = 0.007) and left putamen (R = −0.67, p = 0.003).1
- Resting-state coupling was not the same: GABA-BOLD correlation at rest was limited to visual cortex and cerebellar lobule VI, with Table 2 reporting a peak Rs = 0.8 in the calcarine/cerebellar cluster.1
- Clinical implications stay indirect: The study involved 20 healthy adults and sentence repetition, so it maps a mechanism rather than proving a treatment route for stuttering, aphasia, laryngeal dystonia, or Parkinsonian speech problems.1
GABA (gamma-aminobutyric acid) is the brain’s main inhibitory neurotransmitter, meaning it helps reduce or gate neural firing rather than simply push activity higher. Dopamine is a neuromodulator involved in movement, reward, motivation, and basal ganglia circuit selection; in speech, it has been tied to left-lateralized striatal activity during speaking.2,3
BOLD fMRI measures blood-oxygen-level-dependent activity, an indirect signal related to local neural work and blood flow. PET (positron emission tomography) can measure receptor binding or neurotransmitter-related tracer displacement. Battistella and Simonyan combined [11C]flumazenil PET for GABAA receptor binding, [11C]raclopride PET for striatal D2/D3 dopamine release, and fMRI during overt sentence production.1
20 Adults Completed Flumazenil PET, Raclopride PET, and Speech fMRI
The study enrolled 20 healthy, monolingual, native English-speaking, right-handed adults with no history of neurological, psychiatric, voice, speech, or language problems. Participants averaged 53.2 years old, with SD = 10.1; the sample included 13 women and 7 men.1
The experimental design deliberately separated 3 signals that are often collapsed in casual discussion:
- GABAA receptor binding: [11C]flumazenil PET estimated whole-brain GABAA receptor availability at rest.
- Dopamine release during speech: [11C]raclopride PET estimated striatal D2/D3 tracer displacement during sentence production, a proxy for endogenous dopamine release.
- Speech-related brain activity: fMRI measured BOLD activity while participants listened to and repeated meaningful English sentences.
The speech task used 10 sentences and a sparse-sampling fMRI design, so participants could repeat sentences during a quiet interval before image acquisition. That matters because overt speech creates head-motion and sound artifacts if the scanner records continuously during articulation.
GABA-BOLD Coupling Was Widespread During Speaking
GABAA receptor binding correlated with speech-production BOLD activity across the speech network rather than only in one motor patch.
Positive correlations appeared in the left supplementary motor area, right inferior frontal gyrus, left inferior and superior parietal lobules, right superior temporal gyrus, left putamen, and cerebellum. The strongest tabled peak values reached Rs = 0.9 in the left superior parietal lobule and the right superior temporal gyrus extending to the insula.1
Supplementary motor area (SMA) helps prepare and sequence voluntary movements, including speech-motor plans. The left SMA correlation was Rs = 0.84 with p = 2 x 10−5. Superior temporal cortex supports auditory and speech-sound processing, while inferior parietal regions help map sensory information onto motor commands. Seeing GABA-BOLD coupling across those nodes fits a speech-control system that coordinates listening, planning, and articulation instead of merely triggering mouth movement.
The negative frontal result sharpened the map. Brain activity in the left inferior frontal gyrus pars orbitalis extending to the middle frontal gyrus showed a negative GABA-BOLD correlation, with Rs = −0.74, p = 7 x 10−4 in the results text and a peak Rs = −0.8 in the reported cluster summary.1
Different parts of the speech network may use inhibitory tone differently depending on whether a region is selecting, sequencing, integrating feedback, or suppressing competing activity.
Resting-State GABA Coupling Was Mostly Visual and Cerebellar
Resting-state fMRI told a different, narrower story. GABAA receptor binding correlated with resting-state activity mainly in the bilateral primary visual cortex and left cerebellar lobule VI, with the table listing a calcarine/cerebellar cluster at Rs = 0.8.1
That contrast is the main calibration point. If GABA binding simply reflected a generic baseline property of the whole brain, the resting-state map should have looked more like the speech-production map. Instead, the broader GABA-BOLD relationships appeared when participants were producing speech. The finding is compatible with task-specific inhibitory control: GABAergic tone may help shape the speech network when the system has to turn heard sentences into timed vocal output.
Dopamine-GABA Interaction Concentrated in Left Caudate and Putamen
Earlier work from the same research line showed that speech production induced left-lateralized striatal dopamine release and coupled that release to functional striatal circuits.2 In the 2026 analysis, raclopride PET again anchored dopamine effects in the dorsal striatum.
Speech-related dopamine release included the left anterior caudate (delta RAC BP = −8.6% +/- 12.0), left posterior caudate (delta RAC BP = −9.5% +/- 12.7), left anterior putamen (delta RAC BP = −6.3% +/- 7.9), left posterior putamen (delta RAC BP = −6.3% +/- 10.0), and right anterior caudate (delta RAC BP = −3.9% +/- 5.6).1
Caudate nucleus and putamen are basal ganglia structures involved in action selection, motor learning, and motor execution. During speech, the left caudate and putamen were the places where dopamine, GABA, and BOLD activity statistically met: the left caudate showed R = −0.63, p = 0.007, and the left putamen showed R = −0.67, p = 0.003 for the dopamine x GABA x speech-production BOLD interaction.1
Those interaction values should not be read as “more dopamine equals better speech” or “more GABA equals better speech.” They point to coordination. Dopamine may tune how inhibitory receptor availability relates to speech-related activity inside striatal circuits, especially on the left side where prior work linked dopamine to lateralized speech-network function.2,3
Adjacent Language Evidence Points to Inhibitory Control, Not a GABA Cure
The 2026 paper fills a specific gap because earlier speech-control work had stronger direct evidence for dopamine than for GABA. Simonyan et al. reported left-lateralized speech-induced striatal dopamine release in healthy humans, and Fuertinger et al. later linked dopamine to left-hemispheric lateralization of speech networks.2,3 The new paper adds direct GABAA receptor binding evidence to that dopamine-centered framework.
Language-related GABA findings elsewhere make the direction plausible without making it clinically settled. Krishnamurthy et al. found that resting-state functional connectivity and local GABA concentrations predicted oral reading of real words and pseudo-words.4
Harris et al. reported that language improvements after transcranial direct-current stimulation in primary progressive aphasia were associated with reduced GABA levels.5 Those studies do not test the same sentence-production mechanism, but they support the broader idea that inhibitory tone can shape language-network behavior.
Speech-control models such as DIVA frame speaking as a feedback-control problem: the brain predicts speech sounds and movements, compares them with auditory and somatosensory feedback, and updates motor commands when output drifts.6 GABA and dopamine fit into that framework as circuit regulators, not as simple on/off switches.
Limitations of This PET-fMRI Speech-Control Study
This study is mechanistic and exploratory by design. It strengthens a neurochemical model of normal speech production, but it does not show that changing GABA or dopamine improves speech in patients.
- Healthy sample: the study involved 20 healthy adults, so the findings may not transfer directly to stuttering, aphasia, laryngeal dystonia, Parkinson’s disease, or other speech disorders.
- Receptor specificity: flumazenil PET measures GABAA receptor binding, not all GABAergic signaling or GABAB receptor biology.
- Dopamine scope: raclopride PET is most useful for striatal D2/D3 dopamine receptor systems and does not map cortical dopamine or D1 pathways.
- Timing mismatch: PET and fMRI measure neurophysiology on different timescales, so correlations cannot prove the fast causal sequence of neuromodulation during speech.
- Task scope: sentence repetition is not the same as conversation, prosody, discourse planning, auditory-feedback perturbation, or speech recovery after injury.
Those limits keep the clinical claim disciplined. The paper makes human speech-control biology more specific: inhibitory receptor binding and dopamine release interact with task-linked activity in the left striatum and broader speech network. It does not yet tell clinicians which drug, stimulation protocol, or behavioral therapy should be used for a particular speech disorder.
Questions About GABA, Dopamine, and Speech Control
Did this study find that GABA causes speech production?
No. The study found correlations between GABAA receptor binding and speech-production BOLD activity, plus dopamine-GABA-BOLD interactions in the left striatum. Correlation does not prove that changing GABA would improve or impair speech.
Why is the left striatum important for speech?
The left caudate and putamen are part of basal ganglia loops that help select, sequence, and regulate motor actions. Speech is a learned motor behavior with cognitive and auditory-feedback demands, so left-striatal tuning is plausible and consistent with earlier dopamine speech studies.2,3
Does this explain stuttering or speech disorders?
Not directly. The study used healthy adults and a sentence-repetition task. It gives a mechanistic map that future disorder studies can test, but it does not diagnose stuttering, aphasia, dystonia, or Parkinsonian speech impairment.
Why compare speaking with resting-state activity?
The comparison helps separate speech-specific coupling from generic baseline brain organization. GABA-BOLD links were broad during speaking but limited at rest, which supports a task-linked interpretation.
Could GABA drugs improve speech?
Possibly in selected disorders, but this paper does not test treatment. GABAergic agents and stimulation-related GABA changes have been studied in some speech and language conditions, yet normal speech-production mapping is only an early mechanistic step.5
References
- Battistella G, Simonyan K. Linking GABAergic and dopaminergic neurotransmission: effects on neural activity during human speech control. Human Brain Mapping. 2026;47:e70531. doi:10.1002/hbm.70531
- Simonyan K, Herscovitch P, Horwitz B. Speech-induced striatal dopamine release is left lateralized and coupled to functional striatal circuits in healthy humans: a combined PET, fMRI and DTI study. NeuroImage. 2013;70:21–32. doi:10.1016/j.neuroimage.2012.12.042
- Fuertinger S, Zinn JC, Sharan AD, Hamzei-Sichani F, Simonyan K. Dopamine drives left-hemispheric lateralization of neural networks during human speech. Journal of Comparative Neurology. 2018;526(5):920–931. doi:10.1002/cne.24375
- Krishnamurthy LC, Krishnamurthy V, Crosson B, et al. Strength of resting state functional connectivity and local GABA concentrations predict oral reading of real and pseudo-words. Scientific Reports. 2019;9:11385. doi:10.1038/s41598-019-47889-9
- Harris AD, Wang Z, Ficek B, Webster K, Edden RA, Tsapkini K. Reductions in GABA following a tDCS-language intervention for primary progressive aphasia. Neurobiology of Aging. 2019;79:75–82. doi:10.1016/j.neurobiolaging.2019.03.011
- Tourville JA, Guenther FH. The DIVA model: a neural theory of speech acquisition and production. Language and Cognitive Processes. 2011;26(7):952–981. doi:10.1080/01690960903498424