
People expect life events to drive adolescent wellbeing — conflicts and breakups push it down, friendships and successes push it up. A 2026 longitudinal twin study from Skaug et al. tested this directly in 2,879 Norwegian twins followed from ages 12 to 20 across three waves and found that genetics, not events, explain most of the long-term stability in wellbeing.1
Research Highlights
- 81% of the time-invariant stability in adolescent wellbeing is genetic. The random-intercept component (the part of wellbeing that stays steady across waves) accounted for ~40% of total variance, and 81% of that stable variance was explained by additive genetic factors.1
- Cross-sectional heritability of wellbeing fell from ~48% at age 15 to 26% at age 20. Heritability estimates were a² = 0.48 (95% CI 0.42–0.54) at Wave 1, 0.46 (0.39–0.52) at Wave 2, and 0.26 (0.16–0.36) at Wave 3 — a notable late-adolescence drop in genetic loading on wellbeing.1
- Negative life events correlate with lower wellbeing concurrently, but the effect decays over time. Cross-sectional r = −0.34 to −0.28 at the same wave, dropping to −0.26 to −0.19 across 2 years and −0.22 to −0.12 across 4 years.1
- After accounting for genetics and trait stability, life events explain ~1% of change in wellbeing. The strongest cross-lagged path (negative dependent life events at Wave 1 to wellbeing at Wave 2) was β = −0.11, explaining 1% of variance. The Wave 2 to Wave 3 path was effectively zero and non-significant.1
- Negative dependent life events were themselves 47–55% heritable. Events the adolescent’s own behavior contributed to (peer conflict, school problems, breakups) were under substantial genetic control. The cross-sectional correlation between life events and wellbeing largely reflects the same genetic factors influencing both.1
The Skaug result lands in a literature with a long-running pattern. Lykken and Tellegen’s 1996 paper “Happiness is a stochastic phenomenon” first put a hedonic-setpoint frame around twin findings, arguing that “trying to be happier may be as futile as trying to be taller.”2
That framing went too far — happiness is not biologically fixed — but the kernel was right. Cross-sectional twin estimates of wellbeing heritability have consistently landed in the 30–50% range, and longitudinal designs keep finding that most of the cross-time stability is genetic.3,4
What Skaug et al. add is a careful within-adolescence picture, with enough waves and a strong-enough analytic design (the random intercept cross-lagged panel model, or RI-CLPM) to separate stable trait variance from wave-to-wave fluctuation, and to ask whether life events predict the fluctuation part once trait stability is held constant.
Skaug 2026: 2,879 Twins, Three Waves, From Age 12 to 20
The data come from the Oslo University Adolescent and Young Adult Twin Project, a Norwegian population-based study of all twins born in 1988–1994.1
The included sample:
- 2,879 twins from 1,483 families.
- 1,094 monozygotic (MZ) twins — identical twins who share ~100% of their segregating genes.
- 1,785 dizygotic (DZ) twins — fraternal twins who share ~50% of their segregating genes, like ordinary siblings.
- 56% female.
Three waves of self-report questionnaires were administered ~2 years apart, with mean ages of 15.2, 16.9, and 19.6 years.1
Wellbeing was measured with the Cantril ladder — a 1-to-10 scale where the participant rates their current life from worst possible (1) to best possible (10). It is the most widely used single-item wellbeing measure, with documented reliability and validity in adolescent samples.
Life events were captured with a 38-item checklist covering events in the past year. The events were sorted into three categories:
- Negative dependent (14 events) — events the adolescent’s own behavior plausibly contributed to, like peer conflict, breakups, or trouble at school.
- Negative independent (19 events) — events outside the adolescent’s control, like a parent’s death or family illness.
- Positive dependent (5 events) — positive events shaped by the adolescent’s actions, like making a new friend.
Positive dependent events showed negligible cross-sectional correlation with wellbeing in this sample, so they were dropped from the longitudinal models.
Cross-Sectional Heritability Falls From ~48% at Age 15 to 26% at Age 20
The first analytic step was a trivariate Cholesky decomposition — a structural equation model that splits variance in wellbeing at each wave into three sources:
- A (additive genetic): the share of variance attributable to genetic factors.
- C (shared environment): the share attributable to environmental factors that make twins in the same family more similar (parenting, neighborhood, school).
- E (nonshared environment): the share attributable to environmental factors unique to each twin, plus measurement error.
The C component was dropped from the best-fitting model (no detectable shared-environment contribution), leaving an AE model. Heritability (a²) by wave:
- Wave 1 (mean age 15.2): a² = 0.48 (95% CI 0.42–0.54). MZ twin correlation = 0.50; DZ = 0.21.1
- Wave 2 (mean age 16.9): a² = 0.46 (95% CI 0.39–0.52). MZ = 0.49; DZ = 0.22.1
- Wave 3 (mean age 19.6): a² = 0.26 (95% CI 0.16–0.36). MZ = 0.28; DZ = 0.11.1
The drop at Wave 3 is meaningful. Heritability of wellbeing nearly halved between mid-adolescence and the threshold of young adulthood.
Skaug et al. checked whether within-wave age variation explained the drop — it didn’t. Whatever is happening at the late-adolescence transition, it is genuine narrowing of the genetic contribution to wellbeing.
One plausible interpretation: as adolescents move into independent young adulthood, the environments they construct for themselves become more variable and more consequential. The same genotype lands in a wider range of environments (college, work, partner, city), and that environmental variance shows up as larger E and smaller a².
81% of the Time-Invariant Stability in Wellbeing Is Genetic
The genetically informative RI-CLPM separates each person’s wellbeing into two components:
- A between-person random intercept capturing the stable, across-wave level of wellbeing for that individual.
- Within-person wave-specific deviations from that stable level — the fluctuating part.
The random intercept accounted for about 40% of the total variance at each measurement wave, with the remaining 60% being time-variant fluctuation around the individual’s stable level.1
Of the stable, time-invariant component:
81% of the variance was explained by additive genetic factors. The remaining 19% was nonshared environmental influence (including measurement error).1
In plain English: when an adolescent’s wellbeing stays consistently high or consistently low across years, that consistency is overwhelmingly genetic, not environmental.
This lines up with Lykken and Tellegen’s hedonic-setpoint argument and with Bartels 2015’s twin-meta synthesis showing wellbeing heritability around 36% with strong genetic continuity.2,3
Negative Life Events Correlate With Wellbeing Cross-Sectionally, But Don’t Move It Over Time
The cross-sectional correlations look exactly like what intuition predicts.
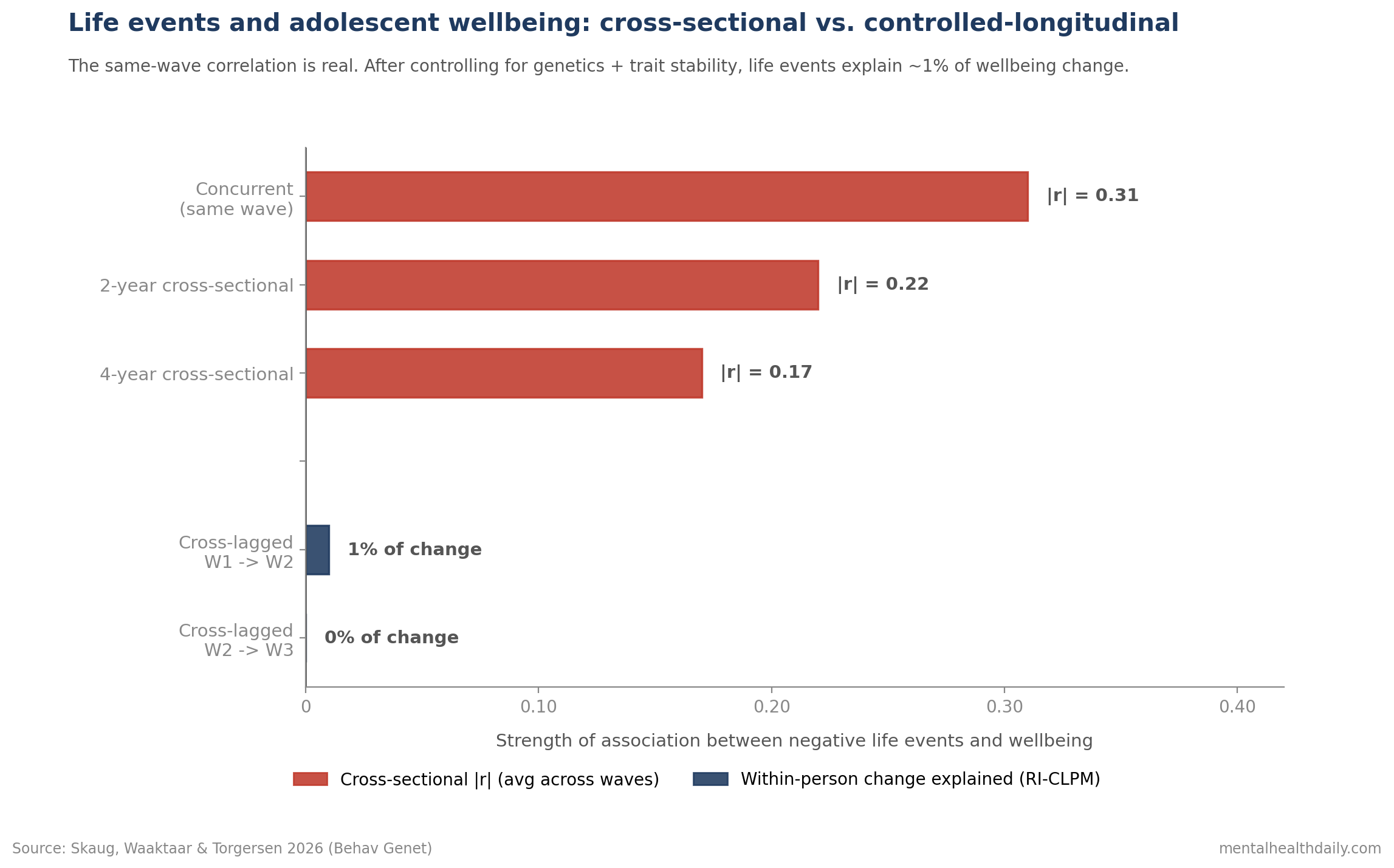
Concurrent correlations are moderate. Life events and wellbeing measured at the same wave correlate r = −0.34 to −0.28. More negative events, lower wellbeing. The pattern holds across the three waves.1
The correlations decay steadily with time. Across 2 years, r dropped to −0.26 to −0.19. Across 4 years, to −0.22 to −0.12.
That decay is the first hint that life events do not lock in long-term wellbeing.
The RI-CLPM cross-lagged paths — which estimate how within-person deviations in life events at one wave predict subsequent within-person changes in wellbeing, after stripping out the stable trait component — are even more striking.
The strongest path: 1% of change explained. Negative dependent life events at Wave 1 predicting wellbeing at Wave 2 came in at β = −0.11, explaining 1% of variance in wellbeing change.1
The Wave 2 to Wave 3 path was effectively zero and non-significant.
The implication is direct. Once you strip out the stable, largely genetic component of wellbeing, the variation in negative life events an adolescent experiences from one wave to the next predicts almost none of the variation in their wellbeing change.
Why Cross-Sectional Correlations Mislead: Gene-Environment Correlation
The story behind the gap between concurrent r = −0.34 and longitudinal effect = 1% is gene-environment correlation (rGE).5
Negative dependent life events in this sample were 47–55% heritable.1
That number deserves a moment of unpacking. It reflects how trait-level dispositions — neuroticism, emotional reactivity, sociability — shape both wellbeing and the life events an adolescent ends up generating, like peer arguments, romantic conflict, or trouble at school.
The same underlying genotype produces:
- Lower baseline wellbeing.
- More negative dependent life events.
So when you cross-sectionally observe that adolescents with more negative events report lower wellbeing, you are partly observing two consequences of the same upstream cause — not the events causing the wellbeing drop.
This is the standard rGE confound, and it is exactly why genetically informed designs like the RI-CLPM exist. Once the design controls for shared genetic influences and stable trait variance, the residual life-event-on-wellbeing effect is the part most plausibly attributable to the events themselves — and that part is small.
Interventions Should Target Trait-Level Skills, Not Event Counts
The Skaug result does not say wellbeing is fixed or that interventions are pointless. It says specific, measurable life events do not, on their own, swing the long-term wellbeing trajectory of adolescents.
- The cross-sectional correlation between life events and wellbeing is real. Adolescents going through a hard period feel worse. The data confirm what clinicians already know.
- The longitudinal effect is small. Most adolescents return toward their stable trait level after a stressful event. This is consistent with adult research on hedonic adaptation and Lyubomirsky’s framework distinguishing setpoint, life circumstances, and intentional activity.6
- Effective interventions probably target the trait, not the events. Cognitive-behavioral skills, regular exercise, social-relationship quality, and meaning-making practices have larger and more durable effects on wellbeing than tweaking the inventory of recent life events. The Lyubomirsky 2005 framework attributes ~40% of wellbeing variance to “intentional activity” — a number that is older and contested, but consistent with the Skaug pattern in the sense that the malleable component is something other than which life events happen to occur.6
- The late-adolescence heritability drop is a window of opportunity. If the genetic loading on wellbeing falls from ~48% to ~26% between ages 15 and 20, the environmental variance is rising. Whatever moves wellbeing in young adulthood has more room to operate than it does at 15.
Norwegian Twin Data Leave Short-Term and Context Limits
It is one population. The Oslo Twin Project is Norwegian, population-based, and has known characteristics. Generalizing to adolescents in lower-income settings or non-Western contexts requires caution.
The wellbeing measure is one item. The Cantril ladder is widely used and has good psychometric properties at the population level, but it asks for a single global judgment. Multidimensional wellbeing measures (positive affect, life satisfaction, eudaimonic dimensions) might show different patterns.
Life-event categorization is imperfect. The dependent/independent split is a useful heuristic but blurs at the edges. “I got into a fight with a friend” reasonably counts as dependent; “my parent died” as independent. Many events sit between.
The model captures average effects, not individual differences in event impact. Some adolescents are devastated by events that others shake off. The Skaug analysis estimates population-average cross-lagged paths; person-level moderators (resilience traits, social support, prior trauma) are not modeled in this paper.
Two-year intervals miss short-term dynamics. Wellbeing fluctuates day-to-day and week-to-week in ways this design cannot detect. Daily-diary or experience-sampling studies show real life-event effects on the timescale of hours and days, even when the 2-year longitudinal effect is small.
Questions About Adolescent Wellbeing Genetics and Life Events
Does this mean parenting and school environment don’t matter for adolescent wellbeing?
No. The shared-environment (C) component was small in this Norwegian sample, but in the Skaug design “shared environment” is a specific quantitative concept — the part of wellbeing variance explained by environments that make twins in the same family similar.
That can be small even when family environments matter for individual development. Family-level variance can be absorbed into the additive-genetic component (when parents pass on both genes and an environment correlated with those genes) or into nonshared environment (when the same family treats two twins differently).
Why does heritability drop at age 20?
The most likely explanation is that the environments young adults construct for themselves — college vs. work, urban vs. rural, partnered vs. single — are more variable than the environments adolescents share with parents and school.
Greater environmental variance means more nonshared environmental contribution to wellbeing variance, which directly lowers the heritability estimate.1
If life events explain 1% of wellbeing change, why do bad events feel so impactful?
Because they really do feel impactful in the short term. The cross-sectional correlation of r = −0.34 means concurrent wellbeing is meaningfully lower during a period of negative events.
What the Skaug analysis adds is that adolescents tend to return toward their stable trait level over the following 2–4 years. The acute effect is real; the persistent effect on the trajectory is small.
Does this support a “genetic setpoint” view of happiness?
Partially. The Lykken & Tellegen 1996 setpoint framing has not held up in its strongest form — happiness is not biologically fixed, and durable changes do occur with sustained behavioral and circumstantial change.2
The calibrated read is that adolescents have a heritable trait-level wellbeing tendency that anchors most long-term variation, but that anchor is not destiny. The 19% of stability variance not explained by genetics, plus the 60% of within-wave variance that is time-variant, leaves substantial room for environment, intervention, and choice to move wellbeing across time.
References
- Stability and change in wellbeing throughout adolescence and its relationship with life events: a longitudinal twin study. Skaug E, Waaktaar T & Torgersen S. Behavior Genetics. 2026. doi:10.1007/s10519-026-10262-4
- Happiness is a stochastic phenomenon. Lykken D & Tellegen A. Psychological Science. 1996;7(3):186-189. doi:10.1111/j.1467-9280.1996.tb00355.x
- Genetics of wellbeing and its components satisfaction with life, happiness, and quality of life: a review and meta-analysis of heritability studies. Bartels M. Behavior Genetics. 2015;45(2):137-156. doi:10.1007/s10519-015-9713-y
- Genetic variants associated with subjective well-being, depressive symptoms, and neuroticism identified through genome-wide analyses. Okbay A et al. Nature Genetics. 2016;48(6):624-633. doi:10.1038/ng.3552
- Genetic influences on measures of the environment: a systematic review. Kendler KS & Baker JH. Psychological Medicine. 2007;37(5):615-626. doi:10.1017/s0033291706009524
- Pursuing happiness: the architecture of sustainable change. Lyubomirsky S, Sheldon KM & Schkade D. Review of General Psychology. 2005;9(2):111-131. doi:10.1037/1089-2680.9.2.111